In molecular crystals the energetic separation between the top of the valence band and the bottom conduction band, i.e. the band gap, is typically 2.5–4 eV, while in inorganic semiconductors
the band gaps are typically 1–2 eV. This implies that molecular
crystals are, in fact, insulators rather than semiconductors in the
conventional sense. They become semiconducting only when charge carriers are either injected from the electrodes or generated by intentional or unintentional doping.
Charge carriers can also be generated in the course of optical
excitation. It is important to realize, however, that the primary
optical excitations are neutral excitons with a Coulomb-binding energy of typically 0.5–1.0 eV. The reason is that in organic semiconductors their dielectric constants
are as low as 3–4. This impedes efficient photogeneration of charge
carriers in neat systems in the bulk. Efficient photogeneration can only
occur in binary systems due to charge transfer between donor and acceptor moieties. Otherwise neutral excitons decay radiatively to the ground state – thereby emitting photoluminescence – or non-radiatively. The optical absorption
edge of organic semiconductors is typically 1.7–3 eV, equivalent to a
spectral range from 700 to 400 nm (which corresponds to the visible
spectrum).
History
Early history
Edge-on view of portion of crystal structure of hexamethyleneTTF/TCNQ charge-transfer salt, highlighting the segregated stacking
In 1862, Henry Letheby obtained a partly conductive material by anodic oxidation of aniline in sulfuric acid. The material was probably polyaniline. In the 1950s, researchers discovered that polycyclic aromatic compounds formed semi-conducting charge-transfer complex salts with halogens. In particular, high conductivity of 0.12 S/cm was reported in perylene–iodinecomplex in 1954. This finding indicated that organic compounds could carry current.
The fact that organic semiconductors are, in principle,
insulators but become semiconducting when charge carriers are injected
from the electrode(s) was discovered by Kallmann and Pope. They found that a hole current can flow through an anthracene
crystal contacted with a positively biased electrolyte containing
iodine that can act as a hole injector. This work was stimulated by the
earlier discovery by Akamatu et al. that aromatic hydrocarbons become conductive when blended with
molecular iodine because a charge-transfer complex is formed. Since it
was readily realized that the crucial parameter that controls injection
is the work function
of the electrode, it was straightforward to replace the electrolyte by a
solid metallic or semiconducting contact with an appropriate work
function. When both electrons and holes are injected from opposite
contacts, they can recombine radiatively and emit light (electroluminescence). It was observed in organic crystals in 1965 by Sano et al.
In 1972, researchers found metallic conductivity in the charge-transfer complex TTF-TCNQ. Superconductivity in charge-transfer complexes was first reported in the Bechgaard salt (TMTSF)2PF6 in 1980.
An organic polymer voltage-controlled switch from 1974. Now in the Smithsonian Chip collection
In 1973 Dr. John McGinness
produced the first device incorporating an organic semiconductor. This
occurred roughly eight years before the next such device was created.
The "melanin (polyacetylenes) bistable switch" currently is part of the chips collection of the Smithsonian Institution.
In 1977, Shirakawa et al. reported high conductivity in oxidized and iodine-doped polyacetylene. They received the 2000 Nobel prize in Chemistry for "The discovery and development of conductive polymers". Similarly, highly conductive polypyrrole was rediscovered in 1979.
Organic LEDs, solar cells and FETs
Rigid-backbone organic semiconductors are now used as active elements in optoelectronic devices such as organic light-emitting diodes (OLED), organic solar cells, organic field-effect transistors
(OFET), electrochemical transistors and recently in biosensing
applications. Organic semiconductors have many advantages, such as easy
fabrication, mechanical flexibility, and low cost.
The discovery by Kallman and Pope paved the way for applying
organic solids as active elements in semiconducting electronic devices,
such as organic light-emitting diodes (OLEDs) that rely on the
recombination of electrons and holes injected from "ohmic" electrodes,
i.e. electrodes with unlimited supply of charge carriers. The next major step towards the technological exploitation of the
phenomenon of electron and hole injection into a non-crystalline organic
semiconductor was the work by Tang and Van Slyke. They showed that efficient electroluminescence can be generated in a
vapor-deposited thin amorphous bilayer of an aromatic diamine (TAPC) and
Alq3 sandwiched between an indium-tin-oxide (ITO) anode and an Mg:Ag
cathode. Another milestone towards the development of organic
light-emitting diodes (OLEDs) was the recognition that also conjugated
polymers can be used as active materials. The efficiency of OLEDs was greatly improved when realizing that phosphorescent states (triplet
excitons) may be used for emission when doping an organic semiconductor
matrix with a phosphorescent dye, such as complexes of iridium with
strong spin–orbit coupling.
Work on conductivity of anthracene crystals contacted with an
electrolyte showed that optically excited dye molecules adsorbed at the
surface of the crystal inject charge carriers. The underlying phenomenon is called sensitized photoconductivity. It
occurs when photo-exciting a dye molecule with appropriate
oxidation/reduction potential adsorbed at the surface or incorporated in
the bulk. This effect revolutionized electrophotography, which is the
technological basis of today's office copying machines. It is also the basis of organic solar cells (OSCs), in which the active element is an electron donor, and an electron acceptor material is combined in a bilayer or a bulk heterojunction.
Doping with strong electron donors or acceptors can render
organic solids conductive even in the absence of light. Examples are
doped polyacetylene and doped light-emitting diodes.
Materials
Amorphous molecular films
Amorphous
molecular films are produced by evaporation or spin-coating. They
have been investigated for device applications such as OLEDs, OFETs, and
OSCs. Illustrative materials are tris(8-hydroxyquinolinato)aluminium, C60, phenyl-C61-butyric acid methyl ester (PCBM), pentacene, carbazoles, and phthalocyanine.
Molecularly doped polymers
Molecularly
doped polymers are prepared by spreading a film of an electrically
inert polymer, e.g. polycarbonate, doped with typically 30% of charge
transporting molecules, on a base electrode. Typical materials are the triphenylenes. They have been investigated for use as photoreceptors in electrophotography. This requires films to have a thickness of several micrometers, which can be prepared using the doctor-blade technique.
Molecular crystals
In
the early days of fundamental research into organic semiconductors the
prototypical materials were free-standing single crystals of the acene
family, e.g. anthracene and tetracene. The advantage of employing molecular crystals instead of amorphous film
is that their charge carrier mobilities are much larger. This is of
particular advantage for OFET applications. Examples are thin films of
crystalline rubrene prepared by hot wall epitaxy.
Neat polymer films
They
are usually processed from solution employing variable deposition
techniques including simple spin-coating, ink-jet deposition or
industrial reel-to-reel coating which allows preparing thin films on a
flexible substrate. The materials of choice are conjugated polymers
such as poly-thiophene, poly-phenylenevinylene, and copolymers of
alternating donor and acceptor units such as members of the
poly(carbazole-dithiophene-benzothiadiazole (PCDTBT) family. For solar cell applications they can be blended with C60 or PCBM as electron acceptors.
Aromatic short peptides self-assemblies
Aromatic short peptides self-assemblies are a kind of promising candidate for bioinspired and durable nanoscale semiconductors. The highly ordered and directional intermolecular π-π interactions
and hydrogen-bonding network allow the formation of quantum confined
structures within the peptide self-assemblies, thus decreasing the band
gaps of the superstructures into semiconductor regions. As a result of the diverse architectures and ease of modification of
peptide self-assemblies, their semiconductivity can be readily tuned,
doped, and functionalized. Therefore, this family of electroactive
supramolecular materials may bridge the gap between the inorganic
semiconductor world and biological systems.
Characterization
Organic
semiconductors can be characterized by UV-photoemission spectroscopy.
The equivalent technique for electron states is inverse photoemission.
To measure the mobility of charge carriers, the traditional technique is the so-called time of flight
(TOF) method. This technique requires relatively thick samples; it is
not applicable to thin films. Alternatively, one can extract the charge
carrier mobility from the current in a field effect transistor as a
function of both the source-drain and the gate voltage. Other ways to
determine the charge carrier mobility involve measuring space charge
limited current (SCLC) flow and "carrier extraction by linearly
increasing voltage (CELIV).
In
contrast to organic crystals investigated in the 1960-70s, organic
semiconductors that are nowadays used as active media in optoelectronic
devices are usually more or less disordered. Combined with the fact that
the structural building blocks are held together by comparatively weak
van der Waals forces this precludes charge transport in delocalized
valence and conduction bands. Instead, charge carriers are localized at
molecular entities, e.g. oligomers or segments of a conjugated polymer
chain, and move by incoherent hopping among adjacent sites with
statistically variable energies. Quite often the site energies feature a
Gaussian distribution. Also the hopping distances can vary
statistically (positional disorder).
A consequence of the energetic broadening of the density of
states (DOS) distribution is that charge motion is both temperature and
field dependent and the charge carrier mobility can be several orders of
magnitude lower than in an equivalent crystalline system. This disorder
effect on charge carrier motion is diminished in organic field-effect
transistors because current flow is confined in a thin layer. Therefore,
the tail states of the DOS distribution are already filled so that the activation energy
for charge carrier hopping is diminished. For this reason the charge
carrier mobility inferred from FET experiments is always higher than
that determined from TOF experiments.
In organic semiconductors, charge carriers couple to vibrational
modes and are referred to as polarons. Therefore, the activation energy
for hopping motion contains an additional term due to structural site
relaxation upon charging a molecular entity. It turns out, however, that
usually the disorder contribution to the temperature dependence of the
mobility dominates over the polaronic contribution.
Mechanical Properties
Elastic Modulus
The
elastic modulus can be measured through tensile testing, which
captures the material's stress-strain response. Additionally, the
buckling method, employing buckling equations and measured wavelengths,
can be used to determine the mechanical modulus of film materials. The elastic modulus significantly impacts the applications of organic
semiconductors; lower moduli are preferable for wearable and flexible
electronics to ensure flexibility, while higher moduli are required for devices needing greater resistance
to mechanical stresses and enhanced structural integrity.
Yield Point
The
yield point of organic semiconductors is the stress or strain level at
which the material starts to deform permanently. After this point, the
material loses its elasticity and undergoes permanent deformation. Yield
strength is usually measured by conducting tensile testing.
Understanding and regulating the yield point of organic semiconductors
is essential to designing devices that can endure operational stress
without permanent deformation. This helps maintain the device's functionality and prolong its lifetime.
Viscoelasticity
As
polymers, organic semiconductors exhibit viscoelasticity, meaning they
exhibit both viscous and elastic characteristics during deformation. Viscoelasticity allows materials to return to their original shape
after being deformed and to exhibit strain that varies over time.
Viscoelasticity is typically measured using dynamic mechanical analysis
(DMA). Viscoelasticity is crucial for wearable devices, which are
subjected to stretching and bending during use. The viscoelastic
properties help the materials absorb energy during these processes,
enhancing durability and ensuring long-term functionality under
continuous physical stress.
A scanning tunneling microscopy image of a single-walled carbon nanotubeRotating single-walled zigzag carbon nanotube
A carbon nanotube (CNT) is a tube made of carbon with a diameter in the nanometre range (nanoscale). They are one of the allotropes of carbon. Two broad classes of carbon nanotubes are recognized:
Single-walled carbon nanotubes (SWCNTs) have diameters around 0.5–2.0 nanometres, about 100,000th the width of a human hair. They can be idealised as cutouts from a two-dimensional graphene sheet rolled up to form a hollow cylinder.
Multi-walled carbon nanotubes (MWCNTs) consist of
nested single-wall carbon nanotubes in a nested, tube-in-tube structure.
Double- and triple-walled carbon nanotubes are special cases of MWCNT.
The predicted properties for SWCNTs were tantalising, but a path to synthesising them was lacking until 1993, when Iijima and Ichihashi at NEC, and Bethune and colleagues at IBM
independently discovered that co-vaporising carbon and transition
metals such as iron and cobalt could specifically catalyse SWCNT
formation. These discoveries triggered research that succeeded in
greatly increasing the efficiency of the catalytic production technique,
and led to an explosion of work to characterise and find applications
for SWCNTs.
The true identity of the discoverers of carbon nanotubes is a subject of some controversy. A 2006 editorial written by Marc Monthioux and Vladimir Kuznetsov in the journal Carbon described the origin of the carbon nanotube. A large percentage of academic and popular literature attributes the
discovery of hollow, nanometre-size tubes composed of graphitic carbon
to Sumio Iijima of NEC
in 1991. His paper initiated a flurry of excitement and could be
credited with inspiring the many scientists now studying applications of
carbon nanotubes. Though Iijima has been given much of the credit for
discovering carbon nanotubes, it turns out that the timeline of carbon
nanotubes goes back much further than 1991.
In 1952, L. V. Radushkevich and V. M. Lukyanovich published clear images of 50-nanometre diameter tubes made of carbon in the Journal of Physical Chemistry Of Russia. This discovery was largely unnoticed, as the article was published in
Russian, and Western scientists' access to Soviet press was limited
during the Cold War. Monthioux and Kuznetsov mentioned in their Carbon editorial:
The
fact is, Radushkevich and Lukyanovich [...] should be credited for the
discovery that carbon filaments could be hollow and have a
nanometre-size diameter, that is to say for the discovery of carbon
nanotubes.
In 1976, Morinobu Endo of CNRS observed hollow tubes of rolled up graphite sheets synthesised by a chemical vapour-growth technique. The first specimens observed would later come to be known as single-walled carbon nanotubes (SWNTs). Endo, in his early review of vapor-phase-grown carbon fibers (VPCF),
also reminded us that he had observed a hollow tube, linearly extended
with parallel carbon layer faces near the fiber core. This appears to be the observation of multi-walled carbon nanotubes at the center of the fiber. The mass-produced MWCNTs today are strongly related to the VPGCF developed by Endo. In fact, they call it the "Endo process", out of respect for his early work and patents. In 1979, John Abrahamson presented evidence of carbon nanotubes at the 14th Biennial Conference of Carbon at Pennsylvania State University.
The conference paper described carbon nanotubes as carbon fibers that
were produced on carbon anodes during arc discharge. A characterization
of these fibers was given, as well as hypotheses for their growth in a
nitrogen atmosphere at low pressures.
In 1981, a group of Soviet scientists published the results of chemical and structural characterization of carbon nanoparticles produced by a thermocatalytic disproportionation of carbon monoxide. Using TEM images and XRD
patterns, the authors suggested that their "carbon multi-layer tubular
crystals" were formed by rolling graphene layers into cylinders. They
speculated that via this rolling, many different arrangements of
graphene hexagonal nets are possible. They suggested two such possible
arrangements: a circular arrangement (armchair nanotube); and a spiral,
helical arrangement (chiral tube).
In 1987, Howard G. Tennent of Hyperion Catalysis was issued a
U.S. patent for the production of "cylindrical discrete carbon fibrils"
with a "constant diameter between about 3.5 and about 70 nanometers...,
length 102 times the diameter, and an outer region of
multiple essentially continuous layers of ordered carbon atoms and a
distinct inner core...."
Helping to create the initial excitement associated with carbon
nanotubes were Iijima's 1991 discovery of multi-walled carbon nanotubes
in the insoluble material of arc-burned graphite rods; and Mintmire, Dunlap, and White's independent prediction that if
single-walled carbon nanotubes could be made, they would exhibit
remarkable conducting properties. Nanotube research accelerated greatly following the independent discoveries by Iijima and Ichihashi at NEC and Bethune et al. at IBM of methods to specifically produce single-walled carbon nanotubes by adding transition-metal catalysts to the carbon in an arc discharge. Thess et al. refined this catalytic method by vaporizing the carbon/transition-metal
combination in a high-temperature furnace, which greatly improved the
yield and purity of the SWNTs and made them widely available for
characterization and application experiments. The arc discharge
technique, well known to produce the famed Buckminsterfullerene, thus played a role in the discoveries of both multi- and single-wall
nanotubes, extending the run of serendipitous discoveries relating to
fullerenes. The discovery of nanotubes remains a contentious issue.
Many believe that Iijima's report in 1991 is of particular importance
because it brought carbon nanotubes into the awareness of the scientific
community as a whole.
In 2020, during an archaeological excavation of Keezhadi in Tamil Nadu, India,
~2600-year-old pottery was discovered whose coatings appear to contain
carbon nanotubes. The robust mechanical properties of the nanotubes are
partially why the coatings have lasted for so many years, say the
scientists.
Structure of SWCNTs
Zigzag nanotube, configuration (8, 0)
Armchair nanotube, configuration (4, 4)
Basic details
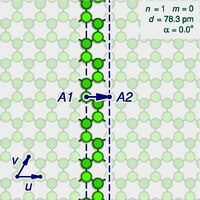
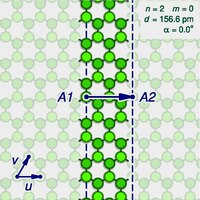
A
"sliced and unrolled" representation of a carbon nanotube as a strip of
a graphene molecule, overlaid on a diagram of the full molecule (faint
background). The arrow shows the gap A2 where the atom A1 on one edge of the strip would fit in the opposite edge, as the strip is rolled upThe basis vectors u and v
of the relevant sub-lattice, the (n,m) pairs that define non-isomorphic
carbon nanotube structures (red dots), and the pairs that define the
enantiomers of the chiral ones (blue dots)
The structure of an ideal (infinitely long) single-walled carbon
nanotube is that of a regular hexagonal lattice drawn on an infinite cylindrical
surface, whose vertices are the positions of the carbon atoms. Since
the length of the carbon-carbon bonds is fairly fixed, there are
constraints on the diameter of the cylinder and the arrangement of the
atoms on it.
In the study of nanotubes, one defines a zigzag path on a graphene-like lattice as a path
that turns 60 degrees, alternating left and right, after stepping
through each bond. It is also conventional to define an armchair path as
one that makes two left turns of 60 degrees followed by two right turns
every four steps. On some carbon nanotubes, there is a closed zigzag
path that goes around the tube. One says that the tube is of the zigzag type or configuration, or simply is a zigzag nanotube. If the tube is instead encircled by a closed armchair path, it is said to be of the armchair type, or an armchair nanotube. An infinite nanotube that is of one type consists entirely of closed paths of that type, connected to each other.
The zigzag and armchair configurations are not the only
structures that a single-walled nanotube can have. To describe the
structure of a general infinitely long tube, one should imagine it being
sliced open by a cut parallel to its axis, that goes through some atom A,
and then unrolled flat on the plane, so that its atoms and bonds
coincide with those of an imaginary graphene sheet—more precisely, with
an infinitely long strip of that sheet. The two halves of the atom A will end up on opposite edges of the strip, over two atoms A1 and A2 of the graphene. The line from A1 to A2 will correspond to the circumference of the cylinder that went through the atom A,
and will be perpendicular to the edges of the strip. In the graphene
lattice, the atoms can be split into two classes, depending on the
directions of their three bonds. Half the atoms have their three bonds
directed the same way, and half have their three bonds rotated 180
degrees relative to the first half. The atoms A1 and A2, which correspond to the same atom A
on the cylinder, must be in the same class. It follows that the
circumference of the tube and the angle of the strip are not arbitrary,
because they are constrained to the lengths and directions of the lines
that connect pairs of graphene atoms in the same class.
Let u and v be two linearly independent vectors that connect the graphene atom A1
to two of its nearest atoms with the same bond directions. That is, if
one numbers consecutive carbons around a graphene cell with C1 to C6,
then u can be the vector from C1 to C3, and v be the vector from C1 to C5. Then, for any other atom A2 with same class as A1, the vector from A1 to A2 can be written as a linear combinationnu + mv, where n and m are integers. And, conversely, each pair of integers (n,m) defines a possible position for A2. Given n and m, one can reverse this theoretical operation by drawing the vector w on the graphene lattice, cutting a strip of the latter along lines perpendicular to w through its endpoints A1 and A2, and rolling the strip into a cylinder so as to bring those two points together. If this construction is applied to a pair (k,0), the result is a zigzag nanotube, with closed zigzag paths of 2k atoms. If it is applied to a pair (k,k), one obtains an armchair tube, with closed armchair paths of 4k atoms.
Types
The structure of the nanotube is not changed if the strip is rotated by 60 degrees clockwise around A1 before applying the hypothetical reconstruction above. Such a rotation changes the corresponding pair (n,m) to the pair (−2m,n+m). It follows that many possible positions of A2 relative to A1 — that is, many pairs (n,m)
— correspond to the same arrangement of atoms on the nanotube. That is
the case, for example, of the six pairs (1,2), (−2,3), (−3,1), (−1,−2),
(2,−3), and (3,−1). In particular, the pairs (k,0) and (0,k) describe the same nanotube geometry. These redundancies can be avoided by considering only pairs (n,m) such that n > 0 and m ≥ 0; that is, where the direction of the vector w lies between those of u (inclusive) and v (exclusive). It can be verified that every nanotube has exactly one pair (n,m) that satisfies those conditions, which is called the tube's type. Conversely, for every type there is a hypothetical nanotube. In fact, two nanotubes have the same type if and only if one can be conceptually rotated and translated so as to match the other exactly. Instead of the type (n,m), the structure of a carbon nanotube can be specified by giving the length of the vector w (that is, the circumference of the nanotube), and the angle α between the directions of u and w,
may range from 0 (inclusive) to 60 degrees clockwise (exclusive). If the diagram is drawn with u horizontal, the latter is the tilt of the strip away from the vertical.
Chiral nanotube of the (3,1) type
Chiral nanotube of the (1,3) type, mirror image of the (3,1) type
Nanotube of the (2,2) type, the narrowest "armchair" one
Nanotube of the (3,0) type, the narrowest "zigzag" one
Chirality and mirror symmetry
A nanotube is chiral if it has type (n,m), with m > 0 and m ≠ n; then its enantiomer (mirror image) has type (m,n), which is different from (n,m). This operation corresponds to mirroring the unrolled strip about the line L through A1 that makes an angle of 30 degrees clockwise from the direction of the u vector (that is, with the direction of the vector u+v). The only types of nanotubes that are achiral are the (k,0) "zigzag" tubes and the (k,k) "armchair" tubes. If two enantiomers are to be considered the same structure, then one may consider only types (n,m) with 0 ≤ m ≤ n and n > 0. Then the angle α between u and w, which may range from 0 to 30 degrees (inclusive both), is called the "chiral angle" of the nanotube.
Circumference and diameter
From n and m one can also compute the circumference c, which is the length of the vector w, which turns out to be:
in picometres. The diameter of the tube is then , that is
also in picometres. (These formulas are only approximate, especially for small n and m where the bonds are strained; and they do not take into account the thickness of the wall.)
The tilt angle α between u and w and the circumference c are related to the type indices n and m by:
where arg(x,y) is the clockwise angle between the X-axis and the vector (x,y); a function that is available in many programming languages as atan2(y,x). Conversely, given c and α, one can get the type (n,m) by the formulas:
which must evaluate to integers.
Physical limits
Narrowest examples
Tube types that are "degenerate" for being too narrow
Degenerate "zigzag" tube type (1,0)
Degenerate "zigzag" tube type (2,0)
Degenerate "armchair" tube type (1,1)
Possibly degenerate chiral tube type (2,1)
If n and m are too small, the structure described by the pair (n,m)
will describe a molecule that cannot be reasonably called a "tube", and
may not even be stable. For example, the structure theoretically
described by the pair (1,0) (the limiting "zigzag" type) would be just a
chain of carbons. That is a real molecule, the carbyne;
which has some characteristics of nanotubes (such as orbital
hybridization, high tensile strength, etc.) — but has no hollow space,
and may not be obtainable as a condensed phase. The pair (2,0) would
theoretically yield a chain of fused 4-cycles; and (1,1), the limiting
"armchair" structure, would yield a chain of bi-connected 4-rings. These
structures may not be realizable.
The thinnest carbon nanotube proper is the armchair structure
with type (2,2), which has a diameter of 0.3 nm. This nanotube was grown
inside a multi-walled carbon nanotube. Assigning of the carbon nanotube
type was done by a combination of high-resolution transmission electron microscopy (HRTEM), Raman spectroscopy, and density functional theory (DFT) calculations.
The thinnest freestanding single-walled carbon nanotube is about 0.43 nm in diameter. Researchers suggested that it can be either (5,1) or (4,2) SWCNT, but
the exact type of the carbon nanotube remains questionable. (3,3), (4,3), and (5,1) carbon nanotubes (all about 0.4 nm in diameter)
were unambiguously identified using aberration-corrected high-resolution transmission electron microscopy inside double-walled CNTs.
Length
Cycloparaphenylene
The observation of the longest carbon nanotubes grown so far, around 0.5 metre (550 mm) long, was reported in 2013. These nanotubes were grown on silicon substrates using an improved chemical vapor deposition (CVD) method and represent electrically uniform arrays of single-walled carbon nanotubes.
The shortest carbon nanotube can be considered to be the organic compound cycloparaphenylene, which was synthesized in 2008 by Ramesh Jasti. Other small molecule carbon nanotubes have been synthesized since.
Density
The highest density of CNTs was achieved in 2013, grown on a conductive titanium-coated copper surface that was coated with co-catalysts cobalt and molybdenum at lower than typical temperatures of 450 °C. The tubes averaged a height of 380 nm and a mass density of 1.6 g cm−3. The material showed ohmic conductivity (lowest resistance ~22 kΩ).
Variants
There
is no consensus on some terms describing carbon nanotubes in the
scientific literature: both "-wall" and "-walled" are being used in
combination with "single", "double", "triple", or "multi", and the
letter C is often omitted in the abbreviation, for example, multi-walled
carbon nanotube (MWNT). The International Standards Organization typically uses "single-walled carbon nanotube (SWCNT)" or "multi-walled carbon nanotube (MWCNT)" in its documents.
Multi-walled
Triple-walled armchair carbon nanotube
Multi-walled nanotubes (MWNTs) consist of multiple rolled layers
(concentric tubes) of graphene. There are two models that can be used to
describe the structures of multi-walled nanotubes. In the Russian Doll model, sheets of graphite
are arranged in concentric cylinders, e.g., a (0,8) single-walled
nanotube (SWNT) within a larger (0,17) single-walled nanotube. In the Parchment
model, a single sheet of graphite is rolled in around itself,
resembling a scroll of parchment or a rolled newspaper. The interlayer
distance in multi-walled nanotubes is close to the distance between
graphene layers in graphite, approximately 3.4 Å. The Russian Doll
structure is observed more commonly. Its individual shells can be
described as SWNTs, which can be metallic or semiconducting. Because of statistical probability and restrictions on the relative
diameters of the individual tubes, one of the shells, and thus the whole
MWNT, is usually a zero-gap metal.
Double-walled carbon nanotubes (DWNTs) form a special class of nanotubes because their morphology and properties are similar to those of SWNTs but they are more resistant to attacks by chemicals. This is especially important when it is necessary to graft chemical functions to the surface of the nanotubes (functionalization) to add properties to the CNT. Covalent functionalization of SWNTs will break some C=C double bonds,
leaving "holes" in the structure on the nanotube and thus modifying
both its mechanical and electrical properties. In the case of DWNTs,
only the outer wall is modified. DWNT synthesis on the gram-scale by the
CCVD technique was first proposed in 2003 from the selective reduction of oxide solutions in methane and hydrogen.
The telescopic motion ability of inner shells, allowing them to
act as low-friction, low-wear nanobearings and nanosprings, may make
them a desirable material in nanoelectromechanical systems (NEMS) . The retraction force that occurs to telescopic motion is caused by the Lennard-Jones interaction between shells, and its value is about 1.5 nN.
Junctions and crosslinking
Transmission electron microscope image of carbon nanotube junction
Junctions between two or more nanotubes have been widely discussed theoretically. Such junctions are quite frequently observed in samples prepared by arc discharge as well as by chemical vapor deposition. The electronic properties of such junctions were first considered theoretically by Lambin et al., who pointed out that a connection between a metallic tube and a
semiconducting one would represent a nanoscale heterojunction. Such a
junction could therefore form a component of a nanotube-based electronic
circuit. The adjacent image shows a junction between two multiwalled
nanotubes.
Junctions between nanotubes and graphene have been considered theoretically and studied experimentally. Nanotube-graphene junctions form the basis of pillared graphene, in which parallel graphene sheets are separated by short nanotubes. Pillared graphene represents a class of three-dimensional carbon nanotube architectures.
3D carbon scaffolds
Recently, several studies have highlighted the prospect of using
carbon nanotubes as building blocks to fabricate three-dimensional
macroscopic (>100 nm in all three dimensions) all-carbon devices.
Lalwani et al. have reported a novel radical-initiated thermal
crosslinking method to fabricate macroscopic, free-standing, porous,
all-carbon scaffolds using single- and multi-walled carbon nanotubes as
building blocks. These scaffolds possess macro-, micro-, and nano-structured pores, and
the porosity can be tailored for specific applications. These 3D
all-carbon scaffolds/architectures may be used for the fabrication of
the next generation of energy storage, supercapacitors, field emission
transistors, high-performance catalysis, photovoltaics, and biomedical
devices, implants, and sensors.
Carbon nanobuds are a newly created material combining two previously discovered allotropes of carbon: carbon nanotubes and fullerenes.
In this new material, fullerene-like "buds" are covalently bonded to
the outer sidewalls of the underlying carbon nanotube. This hybrid material has useful properties of both fullerenes and carbon nanotubes. In particular, they have been found to be exceptionally good field emitters. In composite materials,
the attached fullerene molecules may function as molecular anchors
preventing slipping of the nanotubes, thus improving the composite's
mechanical properties.
A carbon peapod is a novel hybrid carbon material which traps fullerene inside a carbon
nanotube. It can possess interesting magnetic properties with heating
and irradiation. It can also be applied as an oscillator during
theoretical investigations and predictions.
In theory, a nanotorus is a carbon nanotube bent into a torus
(doughnut shape). Nanotori are predicted to have many unique
properties, such as magnetic moments 1000 times larger than that
previously expected for certain specific radii. Properties such as magnetic moment, thermal stability, etc. vary widely depending on the radius of the torus and the radius of the tube.
Graphenated carbon nanotubes are a relatively new hybrid that combines graphitic
foliates grown along the sidewalls of multiwalled or bamboo-style CNTs.
The foliate density can vary as a function of deposition conditions
(e.g., temperature and time) with their structure ranging from a few
layers of graphene (< 10) to thicker, more graphite-like. The fundamental advantage of an integrated graphene-CNT
structure is the high surface area three-dimensional framework of the
CNTs coupled with the high edge density of graphene. Depositing a high
density of graphene foliates along the length of aligned CNTs can
significantly increase the total charge capacity per unit of nominal area as compared to other carbon nanostructures.
Cup-stacked carbon nanotubes (CSCNTs) differ from other quasi-1D
carbon structures, which normally behave as quasi-metallic conductors of
electrons. CSCNTs exhibit semiconducting behavior because of the
stacking microstructure of graphene layers.
Properties
Many properties of single-walled carbon nanotubes depend significantly on the (n,m) type, and this dependence is non-monotonic (see Kataura plot). In particular, the band gap can vary from zero to about 2 eV and the electrical conductivity can show metallic or semiconducting behavior.
Carbon nanotubes are the strongest and stiffest materials yet discovered in terms of tensile strength and elastic modulus. This strength results from the covalent sp2
bonds formed between the individual carbon atoms. In 2000, a
multiwalled carbon nanotube was tested to have a tensile strength of
63 GPa (9,100,000 psi). (For illustration, this translates into the ability to endure tension
of a weight equivalent to 6,422 kilograms-force (62,980 N; 14,160 lbf)
on a cable with cross-section of 1 mm2 (0.0016 sq in)).
Further studies, such as one conducted in 2008, revealed that individual
CNT shells have strengths of up to ≈100 GPa (15,000,000 psi), which is
in agreement with quantum/atomistic models. Because carbon nanotubes have a low density for a solid of 1.3 to 1.4 g/cm3, its specific strength of up to 48,000 kN·m/kg is the best of known materials, compared to high-carbon steel's 154 kN·m/kg.
Although the strength of individual CNT shells is extremely high,
weak shear interactions between adjacent shells and tubes lead to
significant reduction in the effective strength of multiwalled carbon
nanotubes and carbon nanotube bundles down to only a few GPa. This limitation has been recently addressed by applying high-energy
electron irradiation, which crosslinks inner shells and tubes, and
effectively increases the strength of these materials to ≈60 GPa for
multiwalled carbon nanotubes and ≈17 GPa for double-walled carbon nanotube bundles. CNTs are not nearly as strong under compression. Because of their hollow structure and high aspect ratio, they tend to undergo buckling when placed under compressive, torsional, or bending stress.
On the other hand, there is evidence that in the radial direction they are rather soft. The first transmission electron microscope observation of radial elasticity suggested that even van der Waals forces can deform two adjacent nanotubes. Later, nanoindentations with an atomic force microscope
were performed by several groups to quantitatively measure the radial
elasticity of multiwalled carbon nanotubes and tapping/contact mode atomic force microscopy was also performed on single-walled carbon nanotubes. Their high Young's modulus
in the linear direction, of on the order of several GPa (and even up to
an experimentally-measured 1.8 TPa, for nanotubes near 2.4 μm in length), further suggests they may be soft in the radial direction.
Electrical
Band
structures computed using a tight binding approximation for (6,0) CNT
(zigzag, metallic), (10,2) CNT (semiconducting) and (10,10) CNT
(armchair, metallic)
Unlike graphene, which is a two-dimensional semimetal, carbon nanotubes are either metallic or semiconducting along the tubular axis. For a given (n,m) nanotube, if n = m, the nanotube is metallic; if n − m
is a multiple of 3 and n ≠ m, then the nanotube is quasi-metallic with a
very small band gap, otherwise the nanotube is a moderate semiconductor. Thus, all armchair (n = m) nanotubes are metallic, and nanotubes (6,4), (9,1), etc. are semiconducting. Carbon nanotubes are not semimetallic because the degenerate point (the
point where the π [bonding] band meets the π* [anti-bonding] band, at
which the energy goes to zero) is slightly shifted away from the K
point in the Brillouin zone because of the curvature of the tube
surface, causing hybridization between the σ* and π* anti-bonding bands,
modifying the band dispersion.
The rule regarding metallic versus semiconductor behavior has
exceptions because curvature effects in small-diameter tubes can
strongly influence electrical properties. Thus, a (5,0) SWCNT that
should be semiconducting in fact is metallic according to the
calculations. Likewise, zigzag and chiral SWCNTs with small diameters
that should be metallic have a finite gap (armchair nanotubes remain
metallic). In theory, metallic nanotubes can carry an electric current density of 4 billion A/cm2, which is more than 1,000 times greater than those of metals such as copper, where for copper interconnects, current densities are limited by electromigration. Carbon nanotubes are thus being explored as interconnects
and conductivity-enhancing components in composite materials, and many
groups are attempting to commercialize highly conducting electrical wire
assembled from individual carbon nanotubes. There are significant
challenges to be overcome however, such as undesired current saturation
under voltage, and the much more resistive nanotube-to-nanotube junctions and
impurities, all of which lower the electrical conductivity of the
macroscopic nanotube wires by orders of magnitude, as compared to the
conductivity of the individual nanotubes.
Because of its nanoscale cross-section, electrons propagate only
along the tube's axis. As a result, carbon nanotubes are frequently
referred to as one-dimensional conductors. The maximum electrical conductance of a single-walled carbon nanotube is 2G0, where G0 = 2e2/h is the conductance of a single ballistic quantum channel.
Because of the role of the π-electron system in determining the electronic properties of graphene, doping
in carbon nanotubes differs from that of bulk crystalline
semiconductors from the same group of the periodic table (e.g.,
silicon). Graphitic substitution of carbon atoms in the nanotube wall by
boron or nitrogen dopants leads to p-type and n-type behavior, respectively, as would be expected in silicon. However, some non-substitutional (intercalated or adsorbed) dopants introduced into a carbon nanotube, such as alkali metals and electron-rich metallocenes,
result in n-type conduction because they donate electrons to the
π-electron system of the nanotube. By contrast, π-electron acceptors
such as FeCl3 or electron-deficient metallocenes function as
p-type dopants because they draw π-electrons away from the top of the
valence band.
Intrinsic superconductivity has been reported, although other experiments found no evidence of this, leaving the claim a subject of debate.
In 2021, Michael Strano, the Carbon P. Dubbs Professor of
Chemical Engineering at MIT, published department findings on the use of
carbon nanotubes to create an electric current. By immersing the structures in an organic solvent, the liquid drew
electrons out of the carbon particles. Strano was quoted as saying,
"This allows you to do electrochemistry, but with no wires," and represents a significant breakthrough in the technology. Future applications include powering micro- or nanoscale robots, as
well as driving alcohol oxidation reactions, which are important in the
chemicals industry.
Crystallographic defects also affect the tube's electrical
properties. A common result is lowered conductivity through the
defective region of the tube. A defect in metallic armchair-type tubes
(which can conduct electricity) can cause the surrounding region to
become semiconducting, and single monatomic vacancies induce magnetic
properties.
Electromechanical
Semiconducting carbon nanotubes have shown piezoresistive
property when applying mechanical force. The structural deformation
causes a change in the band gap which effects the conductance. This
property has the potential to be used in strain sensors.
Carbon nanotubes have useful absorption, photoluminescence (fluorescence), and Raman spectroscopy
properties. Spectroscopic methods offer the possibility of quick and
non-destructive characterization of relatively large amounts of carbon
nanotubes. There is a strong demand for such characterization from the
industrial point of view: numerous parameters of nanotube synthesis
can be changed, intentionally or unintentionally, to alter the nanotube
quality, such as the non-tubular carbon content, structure (chirality)
of the produced nanotubes, and structural defects. These features then
determine nearly all other significant optical, mechanical, and
electrical properties.
Carbon nanotube optical properties have been explored for use in applications such as for light-emitting diodes (LEDs) and photo-detectors based on a single nanotube have been produced in the lab. Their unique
feature is not the efficiency, which is yet relatively low, but the
narrow selectivity in the wavelength of emission and detection of light and the possibility of its fine-tuning through the nanotube structure. In addition, bolometer and optoelectronic memory devices have been realised on ensembles of single-walled carbon
nanotubes. Nanotube fluorescence has been investigated for the purposes
of imaging and sensing in biomedical applications.
All nanotubes are expected to be very good thermal conductors along the tube, exhibiting a property known as "ballistic conduction",
but good insulators lateral to the tube axis. Measurements show that an
individual SWNT has a room-temperature thermal conductivity along its
axis of about 3500 W·m−1·K−1; compare this to copper, a metal well known for its good thermal conductivity, which transmits 385 W·m−1·K−1. An individual SWNT has a room-temperature thermal conductivity lateral to its axis (in the radial direction) of about 1.52 W·m−1·K−1, which is about as thermally conductive as soil. Macroscopic assemblies
of nanotubes such as films or fibres have reached up to 1500 W·m−1·K−1 so far. Networks composed of nanotubes demonstrate different values of thermal conductivity, from the level of thermal insulation with the thermal conductivity of 0.1 W·m−1·K−1 to such high values. That is dependent on the amount of contribution to the thermal
resistance of the system caused by the presence of impurities,
misalignments and other factors. The temperature stability of carbon
nanotubes is estimated to be up to 2800 °C in vacuum and about 750 °C in air.
Crystallographic defects strongly affect the tube's thermal properties. Such defects lead to phonon scattering, which in turn increases the relaxation rate of the phonons. This reduces the mean free path
and reduces the thermal conductivity of nanotube structures. Phonon
transport simulations indicate that substitutional defects such as
nitrogen or boron will primarily lead to the scattering of
high-frequency optical phonons. However, larger-scale defects such as Stone–Wales defects cause phonon scattering over a wide range of frequencies, leading to a greater reduction in thermal conductivity.
Recently,
carbon-nanotubes have been shown to have antibacterial properties. They
disrupt normal bacterial function by causing physical/mechanical
damage, facilitating oxidative stress
or lipid extraction, inhibiting bacterial metabolism, and isolating
functional sites via wrapping with CNM-containing nanomaterials.
Techniques have been developed to produce nanotubes in sizeable quantities, including arc discharge, laser ablation, chemical vapor deposition
(CVD) and high-pressure carbon monoxide disproportionation (HiPCO).
Among these arc discharge, laser ablation are batch by batch process,
Chemical Vapor Deposition can be used both for batch by batch or
continuous processes, and HiPCO is gas phase continuous process. Most of these processes take place in a vacuum or with process gases.
The CVD growth method is popular, as it yields high quantity and has a
degree of control over diameter, length and morphology. Using
particulate catalysts, large quantities of nanotubes can be synthesized
by these methods, and industrialisation is well on its way, with several
CNT and CNT fibers factory around the world. One problem of CVD
processes is the high variability in the nanotube's characteristics. The HiPCO process advances in catalysis and continuous growth are making CNTs more commercially viable. The HiPCO process helps in producing high purity single-walled carbon
nanotubes in higher quantity. The HiPCO reactor operates at high temperature 900–1100 °C and high pressure ~30–50 bar. It uses carbon monoxide as the carbon source and iron pentacarbonyl or nickel tetracarbonyl as a catalyst. These catalysts provide a nucleation site for the nanotubes to grow, while cheaper iron-based catalysts like Ferrocene can be used for CVD process.
Vertically aligned carbon nanotube arrays
are also grown by thermal chemical vapor deposition. A substrate
(quartz, silicon, stainless steel, carbon fibers, etc.) is coated with a
catalytic metal (Fe, Co, Ni) layer. Typically that layer is iron and is
deposited via sputtering to a thickness of 1–5 nm. A 10–50 nm
underlayer of alumina is often also put down on the substrate first.
This imparts controllable wetting and good interfacial properties.
When the substrate is heated to the growth temperature (~600 to 850 °C),
the continuous iron film breaks up into small islands with each island
then nucleating a carbon nanotube. The sputtered thickness controls the
island size and this in turn determines the nanotube diameter. Thinner
iron layers drive down the diameter of the islands and drive down the
diameter of the nanotubes grown. The amount of time the metal island can
sit at the growth temperature is limited as they are mobile and can
merge into larger (but fewer) islands. Annealing at the growth
temperature reduces the site density (number of CNT/mm2) while increasing the catalyst diameter.
The as-prepared carbon nanotubes always have impurities such as
other forms of carbon (amorphous carbon, fullerene, etc.) and
non-carbonaceous impurities (metal used for catalyst). These impurities need to be removed to make use of the carbon nanotubes in applications.
Purification
As-synthesized
carbon nanotubes typically contain impurities and most importantly
different chiralities of carbon nanotubes. Therefore, multiple methods
have been developed to purify them including polymer-assisted,density gradient ultracentrifugation (DGU), chromatography and aqueous two-phase extraction (ATPE). These methods have been reviewed in multiple articles.
Certain polymers selectively disperse or wrap CNTs of a
particular chirality, metallic character or diameter. For example,
poly(phenylenevinylenes) disperses CNTs of specific diameters
(0.75–0.84 nm) and polyfluorenes are highly selective for semiconducting
CNTs. It involves mainly two steps, sonicate the mixture (CNTs and
polymers in solvent), centrifuge and the supernatant are desired CNTs.
Density gradient ultracentrifugation is a method based on the
density difference of CNTs, so that different components are layered in
centrifuge tubes under centrifugal force. Chromatography-based methods
include size exclusion (SEC), ion-exchange (IEX) and gel chromatography.
For SEC, CNTs are separated due to the difference in size using a
stationary phase with different pore size. As for IEX, the separation is
achieved based on their differential adsorption and desorption
onto chemically functionalized resins packed in an IEX column, so
understanding the interaction between CNTs mixtures and resins is
important. The first IEX is reported to separate DNA-SWCNTs. Gel chromatography is based on the partition of CNTs between stationary
and mobile phase, it's found semiconducting CNTs are more strongly
attracted by gel than metallic CNTs. While it shows potential, the current application is limited to the separation of semiconducting (n,m) species.
ATPE uses two water-soluble polymers such as polyethylene glycol (PEG) and dextran. When mixed, two immiscible
aqueous phases form spontaneously, and each of the two phases shows a
different affinity to CNTs. Partition depends on the solvation energy
difference between two similar phases of microscale volumes. By changing
the separation system or temperatures, and adding strong oxidants,
reductants, or salts, the partition of CNTs species into the two phases
can be adjusted.
Despite the progress that has been made to separate and purify
CNTs, many challenges remain, such as the growth of chirality-controlled
CNTs, so that no further purification is needed, or large-scale
purification.
Advantages of monochiral CNTs
Monochiral
CNTs have the advantage that they do contain less or no impurities,
well-defined non-congested optical spectra. This allows to create for
example CNT-based biosensors with higher sensitivity and selectivity. For example, monochiral SWCNTs are necessary for multiplexed and ratiometric sensing schemes, enhanced sensitivity of biocompatibility.
Carbon nanotubes can be functionalized to attain desired properties that can be used in a wide variety of applications. The two main methods of carbon nanotube functionalization are covalent
and non-covalent modifications. Because of their apparent hydrophobic
nature, carbon nanotubes tend to agglomerate hindering their dispersion in
solvents or viscous polymer melts. The resulting nanotube bundles or
aggregates reduce the mechanical performance of the final composite. The
surface of the carbon nanotubes can be modified to reduce the hydrophobicity and improve interfacial adhesion to a bulk polymer through chemical attachment.
Chemical routes such as covalent functionalization have been
studied extensively, which involves the oxidation of CNTs via strong
acids (e.g. sulfuric acid,
nitric acid, or a mixture of both) in order to set the carboxylic
groups onto the surface of the CNTs as the final product or for further
modification by esterification or amination. Free radical grafting is a
promising technique among covalent functionalization methods, in which
alkyl or aryl peroxides, substituted anilines, and diazonium salts are
used as the starting agents.
Functionalization can improve CNTs characteristically weak
dispersibility in many solvents, such as water - a consequence of their
strong intermolecular p–p interactions. This can enhance the processing
and manipulation of insoluble CNTs, rendering them useful for
synthesizing innovative CNT nanofluids with impressive properties that are tunable for a wide range of applications.
Free radical grafting of macromolecules (as the functional group)
onto the surface of CNTs can improve the solubility of CNTs compared to
common acid treatments which involve the attachment of small molecules
such as hydroxyl onto the surface of CNTs. The solubility of CNTs can be
improved significantly by free-radical grafting because the large
functional molecules facilitate the dispersion of CNTs in a variety of
solvents even at a low degree of functionalization. Recently an
innovative environmentally friendly approach has been developed for the
covalent functionalization of multi-walled carbon nanotubes (MWCNTs)
using clove buds. This approach is innovative and green because it does
not use toxic and hazardous acids which are typically used in common
carbon nanomaterial functionalization procedures. The MWCNTs are
functionalized in one pot using a free radical grafting reaction. The
clove-functionalized MWCNTs are then dispersed in water producing a
highly stable multi-walled carbon nanotube aqueous suspension
(nanofluids).
The surface of carbon nanotubes can be chemically modified by coating spinel nanoparticles by hydrothermal synthesis and can be used for water oxidation purposes.
In addition, the surface of carbon nanotubes can be fluorinated
or halofluorinated by heating while in contact with a fluoroorganic
substance, thereby forming partially fluorinated carbons (so-called
Fluocar materials) with grafted (halo)fluoroalkyl functionality.
Modeling
Computer simulated microstructures with agglomeration regions
Carbon nanotubes are modelled in a similar manner as traditional
composites in which a reinforcement phase is surrounded by a matrix
phase. Ideal models such as cylindrical, hexagonal and square models are
common. The size of the micromechanics
model is highly function of the studied mechanical properties. The
concept of representative volume element (RVE) is used to determine the
appropriate size and configuration of the computer model to replicate
the actual behavior of the CNT-reinforced nanocomposite. Depending on
the material property of interest (thermal, electrical, modulus, creep),
one RVE might predict the property better than the alternatives. While
the implementation of the ideal model is computationally efficient, they
do not represent microstructural features observed in scanning electron
microscopy of actual nanocomposites. To incorporate realistic modeling,
computer models are also generated to incorporate variability such as
waviness, orientation and agglomeration of multiwall or single-wall
carbon nanotubes.
For single-wall carbon nanotubes, ISO/TS 10868 describes a measurement method for the diameter, purity, and fraction of metallic nanotubes through opticalabsorption spectroscopy, while ISO/TS 10797 and ISO/TS 10798 establish methods to characterize
the morphology and elemental composition of single-wall carbon
nanotubes, using transmission electron microscopy and scanning electron microscopy respectively, coupled with energy dispersive X-ray spectrometry analysis.
For multiwall carbon nanotubes, ISO/TR 10929 identifies the basic properties and the content of impurities, while ISO/TS 11888 describes morphology using scanning electron microscopy, transmission electron microscopy, viscometry, and light scattering analysis. ISO/TS 10798 is also valid for multiwall carbon nanotubes.
The National Institute for Occupational Safety and Health
(NIOSH) is the leading United States federal agency conducting research
and providing guidance on the occupational safety and health
implications and applications of nanomaterials.
Early scientific studies have indicated that nanoscale particles may
pose a greater health risk than bulk materials due to a relative
increase in surface area per unit mass. Increase in length and diameter
of CNT is correlated to increased toxicity and pathological alterations in the lungs. The biological interactions of nanotubes are not well understood, and
the field is open to continued toxicological studies. It is often
difficult to separate confounding factors, and since carbon is
relatively biologically inert, some of the toxicity attributed to carbon
nanotubes may be instead due to residual metal catalyst contamination.
In previous studies, only Mitsui-7 was reliably demonstrated to be
carcinogenic, although for unclear/unknown reasons. Unlike many common mineral fibers (such as asbestos), most SWCNTs and
MWCNTs do not fit the size and aspect-ratio criteria to be classified as
respirable fibers. In 2013, given that the long-term health effects
have not yet been measured, NIOSH published a Current Intelligence
Bulletin detailing the potential hazards and recommended exposure limit for carbon nanotubes and fibers. The U.S. National Institute for Occupational Safety and Health has determined non-regulatory recommended exposure limits (RELs) of 1 μg/m3 for carbon nanotubes and carbon nanofibers as background-corrected elemental carbon as an 8-hour time-weighted average (TWA) respirable mass concentration. Although CNT caused pulmonary inflammation and toxicity in mice,
exposure to aerosols generated from sanding of composites containing
polymer-coated MWCNTs, representative of the actual end-product, did not
exert such toxicity.
As of October 2016, single-wall carbon nanotubes have been registered through the European Union's Registration, Evaluation, Authorization and Restriction of Chemicals
(REACH) regulations, based on evaluation of the potentially hazardous
properties of SWCNT. Based on this registration, SWCNT commercialization
is allowed in the EU up to 100 metric tons. Currently, the type of SWCNT registered through REACH is limited to the
specific type of single-wall carbon nanotubes manufactured by OCSiAl, which submitted the application.
Numerous
studies have identified key physicochemical properties of carbon
nanotubes (CNTs) that influence their biological interactions and
toxicological profiles in both in vitro and in vivo systems:
Aspect ratio, length, and rigidity: Long and rigid CNTs
exhibit increased biopersistence and are strongly associated with
fibrogenesis, through the activation of the Smad2/3 and ERK1/2 signaling
pathways. Their high aspect ratio and fiber-like morphology, reminiscent of
asbestos, can impair macrophage-mediated clearance, and induce
inflammation, fibrotic tissue remodeling, granuloma formation, and even DNA damage.
Multi-walled carbon nanotubes (MWCNTs) with lengths ranging from
approximately 0.5 to 10 μm have been implicated in severe pulmonary
conditions, including asbestosis-like fibrosis and mesothelioma. Similarly, single-walled carbon nanotubes (SWCNTs) exceeding 10 μm in
length can induce granulomatous lesions. In contrast, shorter
SWCNTs—particularly those under 300 nm and well-dispersed—are more
readily cleared from the body via renal and biliary excretion pathways. These shorter, purified SWCNTs have demonstrated significantly reduced
inflammatory and cytotoxic responses and, to date, lack conclusive
evidence of carcinogenicity.
Aggregation state: The aggregation state of CNTs refers to
their propensity to cluster into bundles or agglomerates, primarily
driven by van der Waals forces and electrostatic interactions. A
positive correlation has been observed between the degree of aggregation
and cytotoxic potential, with CNT agglomerates exhibiting greater
toxicity than CNT bundles.
Agglomerated CNTs tend to form larger, more rigid, and compact
structures—morphologically similar to asbestos fibers—compared to the
more loosely associated CNT bundles. Specifically, agglomerated SWCNTs have been shown to induce pronounced
granulomatous inflammation, characterized by granuloma formation
surrounded by hypertrophic macrophages in seven days post-exposure. In
contrast, well-dispersed SWCNTs elicit significantly milder inflammatory
responses.
Impurities: Metal contaminants of CNT materials are
strongly associated with the induction of oxidative stress and
proinflammatory responses, with toxicity levels closely correlated to
metal content. Among them, iron is a potent catalyst for intracellular reactive oxygen
species generation, disrupting iron homeostasis and promoting oxidative
damage. Nickel exhibits high bioactivity and proinflammatory potential. The
underlying mechanism is thought to involve lysosomal membrane
destabilization, leading to the release of cathepsin B and subsequent
activation of the NLRP3 inflammasome pathway.
Beyond metal impurities, the presence of non-metallic
contaminants, including amorphous carbon and other carbonaceous
byproducts (e.g., polycyclic aromatic hydrocarbons can significantly
modulate the biological response to CNTs, contributing to cytotoxicity
and inflammatory signaling.
Mitigate carbon nanotube toxicity via physicochemical modulation
Length, diameter, and aggregation state:
As-synthesized CNTs typically form large bundles or agglomerates, which
are associated with increased cytotoxicity. Disaggregation into
individualized CNTs and cutting length can mitigate toxicity. These
modifications are commonly achieved through tip ultrasonication in the
presence of surfactants or polymers, followed by ultracentrifugation to
remove residual bundles and impurities. Such processing enhances
dispersion, reduces rigidity, and improves biocompatibility.
Single-type enrichment and classification: Heterogeneity
in CNT materials contributes to inconsistent biological responses.
Advanced sorting techniques, such as aqueous two-phase extraction, size-exclusion chromatography and density gradient ultracentrifugation, can sort CNTs by specific parameters such as diameter, length, number of walls, and chirality.
Purity enhancement: Post-synthesis purification techniques—such as mild oxidation, sonication in hydrogen peroxide or acetone, acid washing, and incandescent annealing—are employed to remove residual metal catalysts and carbonaceous byproducts.
Surface modification:
Non-covalent modification: Wrapping CNTs with biocompatible polymers such as single-stranded DNA (ssDNA), phospholipid–polyethylene glycol, or Pluronic F108 enhances solubility, colloidal stability, and biological compatibility
without altering the intrinsic structure of the nanotubes.
Covalent modification: Functionalization with carboxyl
groups can reduce cellular and pulmonary toxicity by suppressing NLRP3
inflammasome activation and downregulating pro-inflammatory cytokines
(e.g., IL-1β, TGF-β1, PDGF-AA). PEGylation, the covalent attachment of polyethylene glycol (PEG),
reduces cellular uptake, protein adsorption, oxidative stress, and
immune activation, reducing immune recognition and systemic
inflammation.
Exposure route considerations in CNT toxicology
Pulmonary exposure: Inhalation of airborne CNTs during manufacturing or handling poses the highest risk. CNTs can induce dose-dependent pulmonary toxicity, including granuloma
formation, fibrosis, and inflammation. MWCNTs, due to their fiber-like
structure, are particularly prone to eliciting asbestos-like responses. SWCNTs may translocate systemically but generally exhibit less chronic pulmonary toxicity.
Dermal and consumer exposure: Intact skin provides a
barrier to CNT penetration, though localized cutaneous inflammation has
been reported while current evidence suggests limited systemic health
impact.
Oral and systemic exposure: Oral ingestion of CNTs
typically results in low bioavailability and rapid excretion, with
significant effects observed only at high doses. Systemic exposure via intravenous or intraperitoneal routes is primarily relevant in experimental or therapeutic contexts.
Applications
Nano tape
Carbon nanotubes are currently used in multiple industrial and consumer applications. These include battery components, polymer
composites, to improve the mechanical, thermal and electrical
properties of the bulk product, and as a highly absorptive black paint.
Many other applications are under development, including field effect
transistors for electronics, high-strength fabrics, biosensors for
biomedical and agricultural applications, and many others.
Biomedical Applications
Because
of their relatively large surface area, CNTs are capable of interacting
with a wide variety of therapeutic and diagnostic agents (drugs, genes,
vaccines, antibodies, biosensors, etc.). This can be utilized to assist
in drug delivery directly into cells. In addition, CNTs have recently been used as reinforcements in implants
and scaffolds due to their suitable reaction area, high elastic
modulus, and load transfer capability.
CNTs have been shown to increase the effectiveness of bioactive
coatings for the attachment, proliferation, and differentiation of osteoblasts, and has been used as a bone substitution material.
CNTs may be used as reinforcing materials for chitosan-containing coatings used on implants and medical scaffolds.
Biosensing
SWCNTs
have nanoscale dimensions that fit to the size of biological species.
Due to this size compatibility and their large surface-to-volume ratio,
they are sensitive to changes in their chemical environment. Through covalent and non-covalent surface functionalization, SWCNTs can
be precisely tailored for selective molecular interactions with a
target analyte. The SWCNT represents the transduction unit that converts the
interaction into a signal change (optical or electrical). Due to
continuous progress in the development of detection strategies, there
are numerous examples of the use of SWCNTs as highly sensitive
nanosensors (even down to the single molecule level) for a variety of important biomolecules. Examples include the detection of reactive oxygen and nitrogen species, neurotransmitters, other small molecules, lipids, proteins,sugars, DNA/RNA, enzymes as well as bacteria.
Optical
biosensors with SWCNTs. The functionalization of SWCNTs with
(bio)polymers leads to nanosensors for various molecules. The
interaction with these molecules influences the NIR fluorescence of the
SWCNTs.
The signal change manifests itself in an increase or decrease in the current (electrical) or in a change in the intensity or wavelength of the fluorescence emission (optical). Depending on the type of application, both electrical or optical signal transmission can be advantageous. For sensitive measurement of electronic changes, field-effect transistors
(FET) are often used in which the flow of charges within the SWCNTs is
measured. The FET structures allow easy on-chip integration and can be
parallelized to detect multiple target analytes simultaneously. However, such sensors are more invasive for in vivo applications, as
the entire device has to be inserted into the body. Optical detection
with semiconducting SWCNTs is based on the radiative recombination of
excitons in the near-infrared (NIR) by prior optical (fluorescence) or electrical excitation (electroluminescence).
The emission in the NIR enables detection in the biological
transparency window, where optical sensor applications benefit from
reduced scattering and autofluorescence of biological samples and
consequently a high signal-to-noise ratio. Compared to optical sensors in the UV or visible
range, the penetration depth in biological tissue is also increased. In
addition to the advantage of a contactless readout SWCNTs have
excellent photostability, which enables long-term sensor applications. Furthermore, the nanoscale
size of SWCNTs allows dense coating of surfaces which enables chemical
imaging, e.g. of cellular release processes with high spatial and
temporal resolution. Detection of several target analytes is possible by the spatial arrangement of different SWCNT sensors in arrays or by hyperspectral detection based on monochiral SWCNT sensors that emit at different emission
wavelengths. For fluorescence applications, however, optical filters to
distinguish between excitation and emission and a NIR-sensitive detector
must be used. Standard silicon detectors can also be used if monochiral
SWCNTs (extractable by special purification processes) emitting closer
to the visible range (800 – 900 nm) are used. In order to avoid susceptibility of optical sensors to fluctuating
ambient light, internal references such as SWCNTs that are modified to
be non-responsive or stable NIR emitters can be used. An alternative is to measure fluorescence lifetimes instead of fluorescence intensities. Overall, SWCNTs therefore have
great potential as building blocks for various biosensors.
To render SWCNTs suitable for biosensing, their surface needs to be
modified to ensure colloidal stability and provide a handle for
biological recognition. Therefore, biosensing and surface modifications
(functionalization) are closely related.
Potential future applications include biomedical and environmental applications such as monitoring plant health in agriculture, standoff process control in bioreactors, research/diagnostics of neuronal communication and numerous diseases such as coagulation disorders, diabetes, cancer, microbial and viral infections, testing the efficacy of pharmaceuticals or infection monitoring using smart implants. In industry, SWCNTs are
already used as sensors in the detection of gases and odors in the form
of an electronic nose or in enzyme screening.
Amroy Europe Oy manufactures Hybtonite carbon nano-epoxy resins where carbon nanotubes have been chemically activated to bond to epoxy,
resulting in a composite material that is 20% to 30% stronger than
other composite materials. It has been used for wind turbines, marine
paints and a variety of sports gear such as skis, ice hockey sticks,
baseball bats, hunting arrows, and surfboards.
"Gecko tape" (also called "nano tape") is often commercially sold as double-sided adhesive tape.
It can be used to hang lightweight items such as pictures and
decorative items on smooth walls without punching holes in the wall. The
carbon nanotube arrays comprising the synthetic setae leave no residue after removal and can stay sticky in extreme temperatures.
Applications of nanotubes in development in academia and industry include:
Medical devices: Using single wall carbon nanotubes in medical
devices results in no skin contamination, high flexibility, and
softness, which are crucial for healthcare applications.
Wearable electronics and 5G/6G communication: Electrodes with single
wall carbon nanotubes (SWCNTs) exhibit excellent electrochemical
properties and flexibility.
Bitumen and asphalt: The world's first test section of road pavement
with single wall carbon nanotubes (SWCNTs) showed a 67% increase in
resistance to cracks and ruts, increasing the lifespan of the materials.
Nanocomposites for aviation, automotive, and renewable energy
markets: Modifying resin with just 0.02% single wall carbon nanotubes
(SWCNTs) increases electrical conductivity by 276% without compromising
the mechanical properties of fiber-reinforced polymers, also improving
flexural properties and delaying thermal degradation.
Additive manufacturing: single wall carbon nanotubes (SWCNTs) are
mixed with a suitable printing medium or used as a filler material in
the printing process, creating complex structures with enhanced
mechanical and electrical properties.
Using carbon nanotubes as a scaffold for diverse microfabrication techniques.
Energy dissipation in self-organized nanostructures under the influence of an electric field.
Using carbon nanotubes for environmental monitoring due to their active surface area and their ability to absorb gases.
Jack Andraka used carbon nanotubes in his pancreatic cancer test.
His method of testing won the Intel International Science and
Engineering Fair Gordon E. Moore Award in the spring of 2012.
The Boeing Company has patented the use of carbon nanotubes for structural health monitoring of composites used in aircraft structures. This technology is hoped to
greatly reduce the risk of an in-flight failure caused by structural
degradation of aircraft.
Zyvex Technologies has also built a 54' maritime vessel, the Piranha Unmanned Surface Vessel,
as a technology demonstrator for what is possible using CNT technology.
CNTs help improve the structural performance of the vessel, resulting
in a lightweight 8,000 lb boat that can carry a payload of 15,000 lb
over a range of 2,500 miles.
IMEC is using carbon nanotubes for pellicles in semiconductor lithography.
In tissue engineering, carbon nanotubes have been used as scaffolding for bone growth.
Carbon nanotubes can serve as additives to various structural
materials. For instance, nanotubes form a tiny portion of the
material(s) in some (primarily carbon fiber) baseball bats, golf clubs, car parts, or damascus steel.
IBM expected carbon nanotube transistors to be used on Integrated Circuits by 2020.
SWCNTs have found use in long lasting, faster charged lithium ion batteries; polyamide car parts for e-painting; automotive primers for cost benefits and better aesthetics of topcoats; ESD floors; electrically conductive lining coatings for tanks and pipes; rubber parts with improved heat and oil aging stability; conductive gelcoats for ATEX requirements and tooling conductive gelcoats for increased safety and efficiency; and heating fiber coatings for infrastructure elements.
The strength and flexibility of carbon nanotubes makes them of
potential use in controlling other nanoscale structures, which suggests
they will have an important role in nanotechnology engineering. The highest tensile strength of an individual multi-walled carbon nanotube has been tested to be 63 GPa. Carbon nanotubes were found in Damascus steel from the 17th century, possibly helping to account for the legendary strength of the swords made of it. Recently, several studies have highlighted the prospect of using carbon
nanotubes as building blocks to fabricate three-dimensional macroscopic
(>1mm in all three dimensions) all-carbon devices. Lalwani et al.
have reported a novel radical initiated thermal crosslinking method to
fabricated macroscopic, free-standing, porous, all-carbon scaffolds
using single- and multi-walled carbon nanotubes as building blocks. These scaffolds possess macro-, micro-, and nano- structured pores and
the porosity can be tailored for specific applications. These 3D
all-carbon scaffolds/architectures may be used for the fabrication of
the next generation of energy storage, supercapacitors, field emission
transistors, high-performance catalysis, photovoltaics, and biomedical devices and implants.
CNTs are potential candidates for future via and wire material in nano-scale VLSI circuits. Eliminating electromigration reliability concerns that plague today's Cu interconnects, isolated (single and multi-wall) CNTs can carry current densities in excess of 1000 MA/cm2 without electromigration damage.
Single-walled nanotubes are likely candidates for miniaturizing
electronics. The most basic building block of these systems is an
electric wire, and SWNTs with diameters of an order of a nanometre can
be excellent conductors. One useful application of SWNTs is in the development of the first intermolecular field-effect transistors (FET). The first intermolecular logic gate using SWCNT FETs was made in 2001. A logic gate requires both a p-FET and an n-FET. Because SWNTs are
p-FETs when exposed to oxygen and n-FETs otherwise, it is possible to
expose half of an SWNT to oxygen and protect the other half from it. The
resulting SWNT acts as a not logic gate with both p- and n-type FETs in the same molecule.
Large quantities of pure CNTs can be made into a freestanding
sheet or film by surface-engineered tape-casting (SETC) fabrication
technique which is a scalable method to fabricate flexible and foldable
sheets with superior properties. Another reported form factor is CNT fiber (a.k.a. filament) by wet spinning. The fiber is either directly spun from the synthesis pot or spun from
pre-made dissolved CNTs. Individual fibers can be turned into a yarn. Apart from its strength and flexibility, the main advantage is making an electrically conducting yarn.
The electronic properties of individual CNT fibers (i.e. bundle of
individual CNT) are governed by the two-dimensional structure of CNTs.
The fibers were measured to have a resistivity
only one order of magnitude higher than metallic conductors at 300 K
(27 °C; 80 °F). By further optimizing the CNTs and CNT fibers, CNT
fibers with improved electrical properties could be developed.
CNT-based yarns are suitable for applications in energy and electrochemical water treatment when coated with an ion-exchange membrane. Also, CNT-based yarns could replace copper as a winding material. Pyrhönen et al. (2015) have built a motor using CNT winding.