From Wikipedia, the free encyclopedia
|
| Binary |
1.01101010000010011110… |
| Decimal |
1.4142135623730950488… |
| Hexadecimal |
1.6A09E667F3BCC908B2F… |
| Continued fraction |
 |
The
square root of 2, or the (1/2)th
power of 2, written in mathematics as
√2 or
21⁄2, is the positive
algebraic number that, when multiplied by itself, gives the number
2. Technically, it is called the
principal square root of 2, to distinguish it from the negative number with the same property.
Geometrically the
square root of 2 is the length of a diagonal across a
square with sides of one unit of length; this follows from the
Pythagorean theorem. It was probably the first number known to be
irrational.
The rational approximation of the square root of two,
665,857/470,832, derived from the fourth step in the
Babylonian algorithm starting with
a0 = 1, is too large by approx.
1.6×10−12: its square is
2.0000000000045…
The rational approximation
99/70 (≈ 1.4142857) is frequently used. Despite having a
denominator of only 70, it differs from the correct value by less than
1/10,000 (approx.
+0.72×10−4). Since it is a convergent of the
continued fraction representation of the square root of two, any better rational approximation has a denominator not less than 169, since
239/169 (≈ 1.4142012) is the next convergent with an error of approx.
−0.12×10−4.
The numerical value for the square root of two, truncated to 65
decimal places, is:
- 1.41421356237309504880168872420969807856967187537694807317667973799... (sequence A002193 in the OEIS).
History
Babylonian clay tablet
YBC 7289 with annotations. Besides showing the square root of 2 in
sexagesimal (
1 24 51 10), the tablet also gives an example where one side of the square is 30 and the diagonal then is
42 25 35. The sexagesimal digit 30 can also stand for
0 30 =
1/2, in which case
0 42 25 35 is approximately 0.7071065.
The
Babylonian clay tablet
YBC 7289 (c. 1800–1600 BC) gives an approximation of
√2 in four
sexagesimal figures,
1 24 51 10, which is accurate to about six
decimal digits,
[1] and is the closest possible three-place sexagesimal representation of
√2:

Another early close approximation is given in
ancient Indian mathematical texts, the
Sulbasutras (c. 800–200 BC) as follows:
Increase the length [of the side] by its third and this third by its own fourth less the thirty-fourth part of that fourth.[2] That is,

This approximation is the seventh in a sequence of increasingly accurate approximations based on the sequence of
Pell numbers, which can be derived from the
continued fraction expansion of
√2. Despite having a smaller denominator, it is only slightly less accurate than the Babylonian approximation.
Pythagoreans
discovered that the diagonal of a square is incommensurable with its
side, or in modern language, that the square root of two is
irrational. Little is known with certainty about the time or circumstances of this discovery, but the name of
Hippasus
of Metapontum is often mentioned. For a while, the Pythagoreans treated
as an official secret the discovery that the square root of two is
irrational, and, according to legend, Hippasus was murdered for
divulging it.
[3][4][5] The square root of two is occasionally called "Pythagoras' number" or "Pythagoras' constant", for example by
Conway & Guy (1996).
[6]
Computation algorithms
There are a number of algorithms for approximating
√2,
which in expressions as a ratio of integers or as a decimal can only be
approximated. The most common algorithm for this, one used as a basis
in many computers and calculators, is the Babylonian method
[7] of computing square roots, which is one of many
methods of computing square roots. It goes as follows:
First, pick a guess,
a0 > 0;
the value of the guess affects only how many iterations are required to
reach an approximation of a certain accuracy. Then, using that guess,
iterate through the following
recursive computation:

The more iterations through the algorithm (that is, the more computations performed and the greater "
n"),
the better approximation of the square root of 2 is achieved. Each
iteration approximately doubles the number of correct digits. Starting
with
a0 = 1 the next approximations are
- 3/2 = 1.5
- 17/12 = 1.416...
- 577/408 = 1.414215...
- 665857/470832 = 1.4142135623746...
The value of
√2 was calculated to 137,438,953,444 decimal places by
Yasumasa Kanada's team in 1997. In February 2006 the record for the calculation of
√2 was eclipsed with the use of a home computer. Shigeru Kondo calculated 1
trillion decimal places in 2010.
[8]
For a development of this record, see the table below. Among
mathematical constants with computationally challenging decimal
expansions, only
π has been calculated more precisely.
[9] Such computations aim to check empirically whether such numbers are
normal.
Record progression
This is a table of recent records in calculating digits of
√2 ( 1 trillion = 1012 = 1,000,000,000,000 ).
| Date |
Name |
Number of digits |
| June 28, 2016 |
Ron Watkins |
10 trillion |
| April 3, 2016 |
Ron Watkins |
5 trillion |
| February 9, 2012 |
Alexander Yee |
2 trillion |
| March 22, 2010 |
Shigeru Kondo |
1 trillion= 1012 |
Reference:[10]
Proofs of irrationality
A short proof of the irrationality of
√2 can be obtained from the
rational root theorem, that is, if
p(x) is a
monic polynomial with integer coefficients, then any
rational root of
p(x) is necessarily an integer. Applying this to the polynomial
p(x) = x2 − 2, it follows that
√2 is either an integer or irrational. Because
√2 is not an integer (2 is not a perfect square),
√2
must therefore be irrational. This proof can be generalized to show
that any root of any natural number which is not the square of a natural
number is irrational.
See
quadratic irrational or
infinite descent for a proof that the square root of any non-square natural number is irrational.
Proof by infinite descent
One proof of the number's irrationality is the following proof by
infinite descent. It is also a
proof by contradiction,
also known as an indirect proof, in that the proposition is proved by
assuming that the opposite of the proposition is true and showing that
this assumption is false, thereby implying that the proposition must be
true.
- Assume that √2 is a rational number, meaning that there exists a pair of integers whose ratio is √2.
- If the two integers have a common factor, it can be eliminated using the Euclidean algorithm.
- Then √2 can be written as an irreducible fraction a/b such that a and b are coprime integers (having no common factor).
- It follows that a2/b2 = 2 and a2 = 2b2. ( (a/b)n = an/bn )
- Therefore, a2 is even because it is equal to 2b2. (2b2 is necessarily even because it is 2 times another whole number and multiples of 2 are even.)
- It follows that a must be even (as squares of odd integers are never even).
- Because a is even, there exists an integer k that fulfills: a = 2k.
- Substituting 2k from step 7 for a in the second equation of step 4: 2b2 = (2k)2 is equivalent to 2b2 = 4k2, which is equivalent to b2 = 2k2.
- Because 2k2 is divisible by two and therefore even, and because 2k2 = b2, it follows that b2 is also even which means that b is even.
- By steps 5 and 8 a and b are both even, which contradicts that a/b is irreducible as stated in step 3.
-
- Q.E.D.
Because there is a contradiction, the assumption (1) that
√2 is a rational number must be false. This means that
√2 is not a rational number; i.e.,
√2 is irrational.
This proof was hinted at by
Aristotle, in his
Analytica Priora, §I.23.
[11] It appeared first as a full proof in
Euclid's
Elements, as proposition 117 of Book X. However, since the early 19th century historians have agreed that this proof is an
interpolation and not attributable to Euclid.
[12]
Proof by unique factorization
An alternative proof uses the same approach with the
fundamental theorem of arithmetic which says every integer greater than 1 has a unique factorization into powers of primes.
- Assume that √2 is a rational number. Then there are integers a and b such that a is coprime to b and √2 = a/b. In other words, √2 can be written as an irreducible fraction.
- The value of b cannot be 1 as there is no integer a the square of which is 2.
- There must be a prime p which divides b and which does not divide a, otherwise the fraction would not be irreducible.
- The square of a can be factored as the product of the primes into which a is factored but with each power doubled.
- Therefore, by unique factorization the prime p which divides b, and also its square, cannot divide the square of a.
- Therefore, the square of an irreducible fraction cannot be reduced to an integer.
- Therefore, √2 cannot be a rational number.
This proof can be generalized to show that if an integer is not an exact
kth power of another integer then its
kth root is irrational. For a proof of the same result which does not rely on the fundamental theorem of arithmetic.
Proof by infinite descent, not involving factoring
The following
reductio ad absurdum argument showing the irrationality of
√2 is less well-known. It uses the additional information
2 > √2 > 1 so that
1 > √2 − 1 > 0.
[13]
- Assume that √2 is a rational number. This would mean that there exist positive integers m and n with n ≠ 0 such that m/n = √2. Then m = n√2 and m√2 = 2n.
- We may assume that n is the smallest integer so that n√2 is an integer. That is, that the fraction m/n is in lowest terms.
- Because 1 > √2 − 1 > 0, it follows from (1) that n > n(√2 − 1) = m − n > 0. So n > m − n > 0.
- Also from (1), we have √2 = m/n = m(√2 − 1)/n(√2 − 1) = 2n − m/m − n.
- Thus the fraction m/n for √2, which according to (2) is already in lowest terms, is represented by (4) in yet lower terms (which follows from the result (3)). This is a contradiction, so the assumption that √2 is rational must be false.
This argument may be tightened as follows.
Let
b be the least positive integer for which
√2 is a rational
a/b. Then
b has the property that twice its square is a square, that is,
2b2 = a2. For a contradiction, we show that
a − b is a smaller positive integer with the same property. Multiply the inequalities
1 > √2 − 1 > 0 by
b to show
b > a − b > 0. Now twice the square of
a − b is
2a2 − 4ab + 2b2. Rewrite the first and last terms using b's property to yield
a2 − 4ab + 4b2, which is just the expansion of
(2b − a)2, the promised square. Thus,
√2 can also be written as
(a − b)/(2 b − a). This procedure can be iterated and understood geometrically, as shown below.
Geometric proof
Figure 1. Stanley Tennenbaum's geometric proof of the irrationality of √2.
The immediately preceding argument has a simple geometric formulation attributed by
John Horton Conway to
Stanley Tennenbaum when the latter was a student in the early 1950s
[14] and whose most recent appearance is in an article by Noson Yanofsky in the May–June 2016 issue of
American Scientist.
[15] Given two squares with integer sides respectively
a and
b,
one of which has twice the area of the other, place two copies of the
smaller square in the larger as shown in Figure 1. The square overlap
region in the middle (
(2b − a)2) must equal the sum of the two uncovered squares (
2(a − b)2).
But these squares on the diagonal have positive integer sides that are
smaller than the original squares. Repeating this process we can find
arbitrarily small squares one twice the area of the other, yet both
having positive integer sides, which is impossible since positive
integers cannot be less than 1.
Figure 2. Tom Apostol's geometric proof of the irrationality of sqrt(2).
Another geometric
reductio ad absurdum argument showing that
√2 is irrational appeared in 2000 in the
American Mathematical Monthly.
[16] It is also an example of proof by
infinite descent. It makes use of classic
compass and straightedge
construction, proving the theorem by a method similar to that employed
by ancient Greek geometers. It is essentially the algebraic proof of the
previous section viewed geometrically in yet another way.
Let
△ABC be a right isosceles triangle with hypotenuse length
m and legs
n as shown in Figure 2. By the
Pythagorean theorem,
m/n = √2. Suppose
m and
n are
integers. Let
m:n be a
ratio given in its
lowest terms.
Draw the arcs
BD and
CE with centre
A. Join
DE. It follows that
AB = AD,
AC = AE and the
∠BAC and
∠DAE coincide. Therefore, the triangles
ABC and
ADE are
congruent by
SAS.
Because
∠EBF is a right angle and
∠BEF is half a right angle,
△BEF is also a right isosceles triangle. Hence
BE = m − n implies
BF = m − n. By symmetry,
DF = m − n, and
△FDC is also a right isosceles triangle. It also follows that
FC = n − (m − n) = 2n − m.
Hence we have an even smaller right isosceles triangle, with hypotenuse length
2n − m and legs
m − n. These values are integers even smaller than
m and
n and in the same ratio, contradicting the hypothesis that
m:n is in lowest terms. Therefore,
m and
n cannot be both integers, hence
√2 is irrational.
Pythagorean theorem proof
The square root of 2 is the diagonal of a square with side lengths 1.
This is another proof by contradiction, supposing that
√2 is rational.
- That means that we can make a right isosceles triangle where the side lengths are natural numbers and the legs and the hypotenuse do not share any common factors (except 1).
- Since the legs are equal, so are their squares. So in order for the Pythagorean theorem
to work for this special right triangle, the square of the hypotenuse
has to be an even number (and if we cut it in half once then we have the
area of the square of the leg).
- Recall that the square of an even number is even and the square of
an odd number is odd. So if the square of the hypotenuse is even the
hypotenuse is even as well.
- Remember that a square is a quadrilateral with 2 pairs of parallel sides which are equal in length and has 4 right angles. So both sides of the square of the hypotenuse are even.
- So the square of the hypotenuse of this right triangle can be cut in
half twice and still have integer area. Since we only want to cut it in
half once, then we'll get an even number.
- So the square of the leg is even. Now according to (2) the leg must be even.
- This contradicts our assumption at (1) that the leg and hypotenuse
have no common factors (except 1). Because if they're both even they
share a common factor of 2. So the assumption that √2 was rational has to be false. Or in other words √2 is an irrational number. Q. E. D.
Analytic proof
- Lemma: let α ∈ ℝ+ and p1, p2,… q1, q2,… ∈ ℕ such that |αqn − pn| ≠ 0 for all n ∈ ℕ and
-


- Then α is irrational.
- Proof: suppose α = a/b with a,b ∈ ℕ+.
- For sufficiently big n

- then


- but aqn − bpn is an integer, absurd, then α is irrational.
- Proof: let p1 = q1 = 1 and


- for all n ∈ ℕ.
- By induction,
-

- for all n ∈ ℕ. For n = 1,

- and if this is true for n then it is true for n + 1. In fact



- By application of the lemma, √2 is irrational.
Constructive proof
In
a constructive approach, one distinguishes between on the one hand not
being rational, and on the other hand being irrational (i.e., being
quantifiably apart from every rational), the latter being a stronger
property. Given positive integers
a and
b, because the
valuation (i.e., highest power of 2 dividing a number) of
2b2 is odd, while the valuation of
a2 is even, they must be distinct integers; thus
|2b2 − a2| ≥ 1. Then
[17]

the latter inequality being true because we assume
a/b ≤ 3 − √2 (otherwise the quantitative apartness can be trivially established). This gives a lower bound of
1/3b2 for the difference
|√2 − a/b|, yielding a direct proof of irrationality not relying on the
law of excluded middle; see
Errett Bishop (1985, p. 18). This proof constructively exhibits a discrepancy between
√2 and any rational.
Properties of the square root of two
Angle size and sector
area are the same when the conic radius is
√2. This diagram illustrates the circular and hyperbolic functions based on sector areas
u.
One-half of
√2, also the
reciprocal of
√2, approximately
0.707106781186548, is a common quantity in geometry and
trigonometry because the
unit vector that makes a 45° angle with the axes in a plane has the coordinates

This number satisfies

One interesting property of
√2 is as follows:

since

This is related to the property of
silver ratios.
√2 can also be expressed in terms of the copies of the
imaginary unit i using only the
square root and
arithmetic operations:

if the square root symbol is interpreted suitably for the complex numbers
i and
−i.
√2 is also the only real number other than 1 whose infinite
tetrate (i.e., infinite exponential tower) is equal to its square. In other words: if for
c > 1 we define
x1 = c and
xn+1 = cxn for
n > 1, we will call the limit of
xn as
n → ∞ (if this limit exists)
f(c). Then
√2 is the only number
c > 1 for which
f(c) = c2. Or symbolically:

√2 appears in
Viète's formula for
π:

for
m square roots and only one minus sign.
[18]
Similar in appearance but with a finite number of terms,
√2 appears in various trigonometric constants:
[19]















It is not known whether
√2 is a
normal number, a stronger property than irrationality, but statistical analyses of its
binary expansion are consistent with the hypothesis that it is normal to base two.
[20]
Series and product representations
The identity
cos π/4 = sin π/4 = 1/√2, along with the infinite product representations for the sine and cosine, leads to products such as

and

or equivalently,

The number can also be expressed by taking the
Taylor series of a trigonometric function. For example, the series for
cos π/4 gives

The Taylor series of
√1 + x with
x = 1 and using the
double factorial n!! gives

The convergence of this series can be accelerated with an
Euler transform, producing

It is not known whether
√2 can be represented with a
BBP-type formula. BBP-type formulas are known for
π√2 and
√2ln(1+√2), however.
[21]
The number can be represented by an infinite series of
Egyptian fractions, with denominators defined by 2
nth terms of a
Fibonacci-like recurrence relation a(n)=34a(n-1)-a(n-2), a(0)=0, a(1)=6.
[22]

Continued fraction representation
The square root of 2 and approximations by convergents of continued fractions
The square root of two has the following
continued fraction representation:

The
convergents
formed by truncating this representation form a sequence of fractions
that approximate the square root of two to increasing accuracy, and that
are described by the
Pell numbers
(known as side and diameter numbers to the ancient Greeks because of
their use in approximating the ratio between the sides and diagonal of a
square). The first convergents are:
1/1, 3/2, 7/5, 17/12, 41/29, 99/70, 239/169, 577/408. The convergent
p/q differs from
√2 by almost exactly
1/2q2√2[citation needed] and then the next convergent is
p + 2q/p + q.
Nested square representations
The following nested square expressions converge to

:

Derived constants
The reciprocal of the square root of two (the square root of
1/2) is a widely used constant.

(sequence
A010503 in the
OEIS)
Paper size
The (approximate)
aspect ratio of
paper sizes under
ISO 216 (A4, A0, etc.) is 1:
√2.
This ratio of lengths of the shorter over the longer side guarantees
that cutting a sheet in half along a line parallel to its shorter side
results in the smaller sheets having the same (approximate) ratio as the
original sheet.
Proof:
Let

shorter length and

longer length of the sides of a sheet of paper, with
 as required by ISO 216.
as required by ISO 216.
Let

be the analogue ratio of the halved sheet, then
 .
.
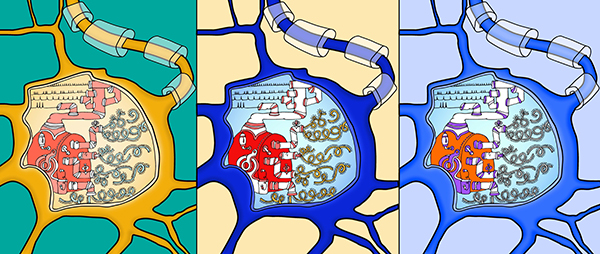 Artist’s
rendition of how the “slow computer” uses geneexpression to process
electrochemical information fromthe “fast computer.” Illustrations:
Anastasia Nizhnik andKelsey Tyssowski
Artist’s
rendition of how the “slow computer” uses geneexpression to process
electrochemical information fromthe “fast computer.” Illustrations:
Anastasia Nizhnik andKelsey Tyssowski
















