Simplified outline of the catabolism of carbohydrates, fatty acids, and amino acids in the synthesis of ATP.
Bioenergetic systems are metabolic processes that relate to the flow of energy in living organisms. Those processes convert energy into adenosine triphosphate (ATP), which is the form suitable for muscular activity. There are two main forms of synthesis of ATP: aerobic, which uses oxygen from the bloodstream, and anaerobic, which does not. Bioenergetics is the field of biology that studies bioenergetic systems.
Overview
The
process that converts the chemical energy of food into ATP (which can
release energy) is not dependent on oxygen availability. During exercise, the supply and demand of oxygen available to muscle cells is affected by duration and intensity and by the individual's cardio respiratory fitness level. It is also affected by the type of activity, for instance, during isometric
activity the contracted muscles restricts blood flow (leaving oxygen
and blood borne fuels unable to be delivered to muscle cells adequately
for oxidative phosphorylation). Three systems can be selectively recruited, depending on the amount of
oxygen available, as part of the cellular respiration process to
generate ATP for the muscles. They are ATP, the anaerobic system and the
aerobic system.
Adenosine triphosphate
ATP is the only type of usable form of chemical energy
for musculoskeletal activity. It is stored in most cells, particularly
in muscle cells. Other forms of chemical energy, such as those available
from oxygen and food, must be transformed into ATP before they can be
utilized by the muscle cells.
Coupled reactions
Since
energy is released when ATP is broken down, energy is required to
rebuild or resynthesize it. The building blocks of ATP synthesis are the
by-products of its breakdown; adenosine diphosphate (ADP) and inorganic phosphate (Pi).
The energy for ATP resynthesis comes from three different series of
chemical reactions that take place within the body. Two of the three
depend upon the food eaten, whereas the other depends upon a chemical
compound called phosphocreatine.
The energy released from any of these three series of reactions is
utilized in reactions that resynthesize ATP. The separate reactions are
functionally linked in such a way that the energy released by one is
used by the other.
Three processes can synthesize ATP:
ATP–CP system (phosphagen system) – At maximum intensity, this system is used for up to 10–15 seconds. The ATP–CP system neither uses oxygen nor produces lactic acid
if oxygen is unavailable and is thus called alactic anaerobic. This is
the primary system behind very short, powerful movements like a golf
swing, a 100 m sprint or powerlifting.
Anaerobic system – This system predominates in supplying energy for
intense exercise lasting less than two minutes. It is also known as the glycolytic system. An example of an activity of the intensity and duration that this system works under would be a 400 m sprint.
Aerobic system – This is the long-duration energy system. After five minutes of exercise, the O2 system is dominant. In a 1 km run, this system is already providing approximately half the energy; in a marathon run it provides 98% or more.
Around mile 20 of a marathon, runners typically "hit the wall," having
depleted their glycogen reserves they then attain "second wind" which is
entirely aerobic metabolism primarily by free fatty acids.
Relative contribution of ATP production of bioenergetic systems during aerobic exercise at maximum intensity (e.g. sprinting)
Aerobic and anaerobic systems usually work concurrently. When
describing activity, it is not a question of which energy system is
working, but which predominates.
Exercise intensity (%Wmax) and substrate use in muscle during aerobic activity (cycling)
The
term aerobic refers to the various series of chemical reactions that
take place within the body. Aerobic refers to the presence of oxygen,
whereas anaerobic means with a series of chemical reactions that does
not require the presence of oxygen. The ATP-CP series and the lactic
acid series are anaerobic, whereas the oxygen series is aerobic.
Anaerobic metabolism
ATP–CP: the phosphagen system
(A)
Phosphocreatine, which is stored in muscle cells, contains a high
energy bond. (B) When creatine phosphate is broken down during muscular
contraction, energy is released and utilized to resynthesize ATP.
Creatine phosphate
(CP), like ATP, is stored in muscle cells. When it is broken down, a
considerable amount of energy is released. The energy released is
coupled to the energy requirement necessary for the resynthesis of ATP.
The total muscular stores of both ATP and CP are small. Thus, the
amount of energy obtainable through this system is limited. The
phosphagen stored in the working muscles is typically exhausted in
seconds of vigorous activity. However, the usefulness of the ATP-CP system lies in the rapid availability of energy rather than quantity. This is important with respect to the kinds of physical activities that humans are capable of performing.
When the phosphagen system has been depleted of phosphocreatine
(creatine phosphate), the resulting AMP produced from the adenylate
kinase (myokinase) reaction is primarily regulated by the purine nucleotide cycle.
Anaerobic glycolysis
The conversion of pyruvate into lactate produces NAD+ to keep glycolysis going
This system is known as anaerobic glycolysis. "Glycolysis"
refers to the breakdown of sugar. In this system, the breakdown of
sugar supplies the necessary energy from which ATP is manufactured. When
sugar is metabolized anaerobically, it is only partially broken down
and one of the byproducts is lactic acid. This process creates enough energy to couple with the energy requirements to resynthesize ATP.
How common monosaccharides (simple sugars) such as glucose, fructose, galactose, and mannose enter the glycolytic pathway
When H+ ions accumulate in the muscles causing the blood pH level to reach low levels, temporary muscle fatigue
results. Another limitation of the lactic acid system that relates to
its anaerobic quality is that only a few moles of ATP can be
resynthesized from the breakdown of sugar. This system cannot be relied
on for extended periods of time.
The lactic acid system, like the ATP-CP system, is important
primarily because it provides a rapid supply of ATP energy. For example,
exercises that are performed at maximum rates for between 1 and 3
minutes depend heavily upon the lactic acid system.
In activities such as running 1500 meters or a mile, the lactic acid
system is used predominantly for the "kick" at the end of the race.
Aerobic metabolism
Aerobic glycolysis
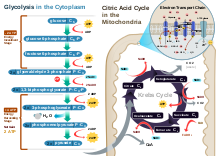
Aerobic glycolysisGlycolysis – The first stage is known as glycolysis, which produces 2 ATP molecules, 2 reduced molecules of nicotinamide adenine dinucleotide (NADH) and 2 pyruvate molecules that move on to the next stage – the Krebs cycle. Glycolysis takes place in the cytoplasm of normal body cells, or the sarcoplasm of muscle cells.
The Krebs cycle – This is the second stage, and the products of this
stage of the aerobic system are a net production of one ATP, one carbon dioxide molecule, three reduced NAD+ molecules, and one reduced flavin adenine dinucleotide (FAD) molecule. (The molecules of NAD+ and FAD mentioned here are electron carriers, and if they are reduced, they have had one or two H+ ions and two electrons added to them.) The metabolites
are for each turn of the Krebs cycle. The Krebs cycle turns twice for
each six-carbon molecule of glucose that passes through the aerobic
system – as two three-carbon pyruvate molecules enter the Krebs cycle. Before pyruvate enters the Krebs cycle it must be converted to acetyl coenzyme A. During this link reaction, for each molecule of pyruvate converted to acetyl coenzyme A, a NAD+ is also reduced. This stage of the aerobic system takes place in the matrix of the cells' mitochondria.
Oxidative phosphorylation – The last stage of the aerobic system
produces the largest yield of ATP – a total of 34 ATP molecules. It is
called oxidative phosphorylation because oxygen is the final acceptor of electrons and hydrogen ions (hence oxidative) and an extra phosphate is added to ADP to form ATP (hence phosphorylation).
This stage of the aerobic system occurs on the cristae
(infoldings of the membrane of the mitochondria). The reaction of each
NADH in this electron transport chain provides enough energy for 3
molecules of ATP, while reaction of FADH2 yields 2 molecules of ATP. This means that 10 total NADH molecules allow the regeneration of 30 ATP, and 2 FADH2
molecules allow for 4 ATP molecules to be regenerated (in total 34 ATP
from oxidative phosphorylation, plus 4 from the previous two stages,
producing a total of 38 ATP in the aerobic system). NADH and FADH2 are oxidized to allow the NAD+
and FAD to be reused in the aerobic system, while electrons and
hydrogen ions are accepted by oxygen to produce water, a harmless
byproduct.
Triglycerides stored in adipose tissue and in other tissues, such as
muscle and liver, release fatty acids and glycerol in a process known as
lipolysis.
Fatty acids are slower than glucose to convert into acetyl-CoA, as
first it has to go through beta oxidation. It takes about 10 minutes for
fatty acids to sufficiently produce ATP. Fatty acids are the primary fuel source at rest and in low to moderate intensity exercise.
Though slower than glucose, its yield is much higher. One molecule of
glucose produces through aerobic glycolysis a net of 30-32 ATP;
whereas a fatty acid can produce through beta oxidation a net of
approximately 100 ATP depending on the type of fatty acid. For example,
palmitic acid can produce a net of 106 ATP.
Normally, amino acids do not provide the bulk of fuel substrates.
However, in times of glycolytic or ATP crisis, amino acids can convert
into pyruvate, acetyl-CoA, and citric acid cycle intermediates.
This is useful during strenuous exercise or starvation as it provides
faster ATP than fatty acids; however, it comes at the expense of risking
protein catabolism (such as the breakdown of muscle tissue) to maintain the free amino acid pool.
The purine nucleotide cycle is used in times of glycolytic or ATP crisis, such as strenuous exercise or starvation. It produces fumarate,
a citric acid cycle intermediate, which enters the mitochondrion
through the malate-aspartate shuttle, and from there produces ATP by
oxidative phosphorylation.
Ketolysis
During starvation or while consuming a low-carb/ketogenic diet, the liver produces ketones. Ketones
are needed as fatty acids cannot pass the blood-brain barrier, blood
glucose levels are low and glycogen reserves depleted. Ketones also
convert to acetyl-CoA faster than fatty acids. After the ketones convert to acetyl-CoA in a process known as ketolysis, it enters the citric acid cycle to produce ATP by oxidative phosphorylation.
The longer that the person's glycogen reserves have been
depleted, the higher the blood concentration of ketones, typically due
to starvation or a low carb diet (βHB 3 - 5 mM). Prolonged aerobic
exercise, where individuals "hit the wall" can create post-exercise ketosis; however, the level of ketones produced are smaller (βHB 0.3 - 2 mM).
Ethanol (alcohol) is first converted into acetaldehyde, consuming NAD+
twice, before being converted into acetate. The acetate is then
converted into acetyl-CoA. When alcohol is consumed in small quantities,
the NADH/NAD+ ratio remains in balance enough for the
acetyl-CoA to be used by the Krebs cycle for oxidative phosphorylation.
However, even moderate amounts of alcohol (1-2 drinks) results in more
NADH than NAD+, which inhibits oxidative phosphorylation.
When the NADH/NAD+ ratio is disrupted (far more NADH than NAD+), this is called pseudohypoxia. The Krebs cycle needs NAD+ as well as oxygen, for oxidative phosphorylation. Without sufficient NAD+,
the impaired aerobic metabolism mimics hypoxia (insufficient oxygen),
resulting in excessive use of anaerobic glycolysis and a disrupted
pyruvate/lactate ratio (low pyruvate, high lactate). The conversion of pyruvate into lactate produces NAD+,
but only enough to maintain anaerobic glycolysis. In chronic excessive
alcohol consumption (alcoholism), the microsomal ethanol oxidizing
system (MEOS) is used in addition to alcohol dehydrogenase.
3D rendering showing thick myocardium within the heart wall.Differently oriented cardiac muscle fibers.Cardiac muscleCardiac sarcomere structure
Cardiac muscle tissue or myocardium forms the bulk of the heart. The
heart wall is a three-layered structure with a thick layer of myocardium
sandwiched between the inner endocardium and the outer epicardium (also known as the visceral pericardium). The inner endocardium lines the cardiac chambers, covers the cardiac valves, and joins with the endothelium that lines the blood vessels that connect to the heart. On the outer aspect of the myocardium is the epicardium which forms part of the pericardial sac that surrounds, protects, and lubricates the heart.
Within the myocardium, there are several sheets of cardiac muscle
cells or cardiomyocytes. The sheets of muscle that wrap around the
left ventricle closest to the endocardium are oriented perpendicularly
to those closest to the epicardium. When these sheets contract in a
coordinated manner they allow the ventricle to squeeze in several
directions simultaneously – longitudinally (becoming shorter from apex
to base), radially (becoming narrower from side to side), and with a
twisting motion (similar to wringing out a damp cloth) to squeeze the
maximum possible amount of blood out of the heart with each heartbeat.
Contracting heart muscle uses a lot of energy, and therefore requires a constant flow of blood to provide oxygen and nutrients. Blood is brought to the myocardium by the coronary arteries. These originate from the aortic root and lie on the outer or epicardial surface of the heart. Blood is then drained away by the coronary veins into the right atrium.
Microanatomy
Illustration of a cardiac muscle cell.
Cardiac muscle cells also called cardiomyocytes are the contractile myocytes of the cardiac muscle. The cells are surrounded by an extracellular matrix produced by supporting fibroblast cells. Specialised modified cardiomyocytes known as pacemaker cells,
set the rhythm of the heart contractions. The pacemaker cells are only
weakly contractile without sarcomeres, and are connected to neighboring
contractile cells via gap junctions. They are located in the sinoatrial node (the primary pacemaker) positioned on the wall of the right atrium, near the entrance of the superior vena cava. Other pacemaker cells are found in the atrioventricular node (secondary pacemaker).
Pacemaker cells carry the impulses that are responsible for the
beating of the heart. They are distributed throughout the heart and are
responsible for several functions. First, they are responsible for
being able to spontaneously generate and send out electrical impulses.
They also must be able to receive and respond to electrical impulses
from the brain. Lastly, they must be able to transfer electrical
impulses from cell to cell.
Pacemaker cells in the sinoatrial node, and atrioventricular node are
smaller and conduct at a relatively slow rate between the cells.
Specialized conductive cells in the bundle of His, and the Purkinje fibers are larger in diameter and conduct signals at a fast rate.
The Purkinje fibers rapidly conduct electrical signals; coronary arteries to bring nutrients to the muscle cells, and veins and a capillary network to take away waste products.
Cardiac muscle cells are the contracting cells that allow the
heart to pump. Each cardiomyocyte needs to contract in coordination
with its neighboring cells - known as a functional syncytium
- working to efficiently pump blood from the heart, and if this
coordination breaks down then – despite individual cells contracting –
the heart may not pump at all, such as may occur during abnormal heart rhythms such as ventricular fibrillation.
Viewed through a microscope, cardiac muscle cells are roughly rectangular, measuring 100–150μm by 30–40μm. Individual cardiac muscle cells are joined at their ends by intercalated discs to form long fibers. Each cell contains myofibrils, specialized protein contractile fibers of actin and myosin that slide past each other. These are organized into sarcomeres,
the fundamental contractile units of muscle cells. The regular
organization of myofibrils into sarcomeres gives cardiac muscle cells a
striped or striated appearance when looked at through a microscope, similar to skeletal muscle. These striations are caused by lighter I bands composed mainly of actin, and darker A bands composed mainly of myosin.
Cardiomyocytes contain T-tubules, pouches of cell membrane
that run from the cell surface to the cell's interior which help to
improve the efficiency of contraction. The majority of these cells
contain only one nucleus (some may have two central nuclei), unlike skeletal muscle cells which contain many nuclei. Cardiac muscle cells contain many mitochondria which provide the energy needed for the cell in the form of adenosine triphosphate (ATP), making them highly resistant to fatigue.
T-tubules
are microscopic tubes that run from the cell surface to deep within the
cell. They are continuous with the cell membrane, are composed of the
same phospholipid bilayer, and are open at the cell surface to the extracellular fluid that surrounds the cell. T-tubules in cardiac muscle are bigger and wider than those in skeletal muscle, but fewer in number.
In the centre of the cell they join, running into and along the cell as
a transverse-axial network. Inside the cell they lie close to the
cell's internal calcium store, the sarcoplasmic reticulum. Here, a single tubule pairs with part of the sarcoplasmic reticulum called a terminal cisterna in a combination known as a diad.
The functions of T-tubules include rapidly transmitting electrical impulses known as action potentials
from the cell surface to the cell's core, and helping to regulate the
concentration of calcium within the cell in a process known as excitation-contraction coupling. They are also involved in mechano-electric feedback, as evident from cell contraction induced T-tubular content exchange (advection-assisted diffusion), which was confirmed by confocal and 3D electron tomography observations.
The cardiac syncytium is a network of cardiomyocytes connected by intercalated discs
that enable the rapid transmission of electrical impulses through the
network, enabling the syncytium to act in a coordinated contraction of
the myocardium. There is an atrial syncytium and a ventricular syncytium that are connected by cardiac connection fibres.
Electrical resistance through intercalated discs is very low, thus
allowing free diffusion of ions. The ease of ion movement along cardiac
muscle fibers axes is such that action potentials are able to travel
from one cardiac muscle cell to the next, facing only slight resistance.
Each syncytium obeys the all or none law.
Intercalated discs are complex adhering structures that connect the single cardiomyocytes to an electrochemical syncytium (in contrast to the skeletal muscle, which becomes a multicellular syncytium during embryonic development).
The discs are responsible mainly for force transmission during muscle
contraction. Intercalated discs consist of three different types of
cell-cell junctions: the actin filament anchoring fascia adherens junctions, the intermediate filament anchoring desmosomes, and gap junctions.
They allow action potentials to spread between cardiac cells by
permitting the passage of ions between cells, producing depolarization
of the heart muscle. The three types of junction act together as a
single area composita.
Under light microscopy,
intercalated discs appear as thin, typically dark-staining lines
dividing adjacent cardiac muscle cells. The intercalated discs run
perpendicular to the direction of muscle fibers. Under electron
microscopy, an intercalated disc's path appears more complex. At low
magnification, this may appear as a convoluted electron dense structure
overlying the location of the obscured Z-line. At high magnification,
the intercalated disc's path appears even more convoluted, with both
longitudinal and transverse areas appearing in longitudinal section.
Cardiac fibroblasts are vital supporting cells within cardiac muscle. They are unable to provide forceful contractions like cardiomyocytes, but instead are largely responsible for creating and maintaining the extracellular matrix which surrounds the cardiomyocytes. Fibroblasts play a crucial role in responding to injury, such as a myocardial infarction. Following injury, fibroblasts can become activated and turn into myofibroblasts – cells which exhibit behaviour somewhere between a fibroblast (generating extracellular matrix) and a smooth muscle cell
(ability to contract). In this capacity, fibroblasts can repair an
injury by creating collagen while gently contracting to pull the edges
of the injured area together.
Fibroblasts are smaller but more numerous than cardiomyocytes,
and several fibroblasts can be attached to a cardiomyocyte at once.
When attached to a cardiomyocyte they can influence the electrical
currents passing across the muscle cell's surface membrane, and in the
context are referred to as being electrically coupled, as originally shown in vitro in the 1960s, and ultimately confirmed in native cardiac tissue with the help of optogenetic techniques. Other potential roles for fibroblasts include electrical insulation of the cardiac conduction system, and the ability to transform into other cell types including cardiomyocytes and adipocytes.
The extracellular matrix (ECM) surrounds the cardiomyocyte and fibroblasts. The ECM is composed of proteins including collagen and elastin along with polysaccharides (sugar chains) known as glycosaminoglycans.
Together, these substances give support and strength to the muscle
cells, create elasticity in cardiac muscle, and keep the muscle cells
hydrated by binding water molecules.
Humans
are born with a set number of heart muscle cells, or cardiomyocytes,
which increase in size as the heart grows larger during childhood
development. Evidence suggests that cardiomyocytes are slowly turned
over during aging, but less than 50% of the cardiomyocytes present at
birth are replaced during a normal life span.
The growth of individual cardiomyocytes not only occurs during normal
heart development, it also occurs in response to extensive exercise (athletic heart syndrome),
heart disease, or heart muscle injury such as after a myocardial
infarction. A healthy adult cardiomyocyte has a cylindrical shape that
is approximately 100μm long and 10–25μm in diameter. Cardiomyocyte
hypertrophy occurs through sarcomerogenesis, the creation of new
sarcomere units in the cell. During heart volume overload,
cardiomyocytes grow through eccentric hypertrophy.
The cardiomyocytes extend lengthwise but have the same diameter,
resulting in ventricular dilation. During heart pressure overload,
cardiomyocytes grow through concentric hypertrophy. The cardiomyocytes grow larger in diameter but have the same length, resulting in heart wall thickening.
The physiology of cardiac muscle shares many similarities with that of skeletal muscle.
The primary function of both muscle types is to contract, and in both
cases, a contraction begins with a characteristic flow of ions across the cell membrane known as an action potential. The cardiac action potential subsequently triggers muscle contraction by increasing the concentration of calcium within the cytosol.
Cardiac cycle
The cardiac cycle
is the performance of the human heart from the beginning of one
heartbeat to the beginning of the next. It consists of two periods: one
during which the heart muscle relaxes and refills with blood, called diastole, following a period of robust contraction and pumping of blood, dubbed systole.
After emptying, the heart immediately relaxes and expands to receive
another influx of blood returning from the lungs and other systems of
the body, before again contracting to pump blood to the lungs and those
systems. A normally performing heart must be fully expanded before it
can efficiently pump again.
The rest phase is considered polarized. The resting potential
during this phase of the beat separates the ions such as sodium,
potassium, and calcium. Myocardial cells possess the property of
automaticity or spontaneous depolarization.
This is the direct result of a membrane which allows sodium ions to
slowly enter the cell until the threshold is reached for depolarization.
Calcium ions follow and extend the depolarization even further. Once
calcium stops moving inward, potassium ions move out slowly to produce
repolarization. The very slow repolarization of the CMC membrane is
responsible for the long refractory period.
However, the mechanism by which calcium concentrations within the
cytosol rise differ between skeletal and cardiac muscle. In cardiac
muscle, the action potential comprises an inward flow of both sodium and
calcium ions. The flow of sodium ions is rapid but very short-lived,
while the flow of calcium is sustained and gives the plateau phase
characteristic of cardiac muscle action potentials. The comparatively
small flow of calcium through the L-type calcium channels triggers a much larger release of calcium from the sarcoplasmic reticulum in a phenomenon known as calcium-induced calcium release.
In contrast, in skeletal muscle, minimal calcium flows into the cell
during action potential and instead the sarcoplasmic reticulum in these
cells is directly coupled to the surface membrane. This difference can
be illustrated by the observation that cardiac muscle fibers require
calcium to be present in the solution surrounding the cell to contract,
while skeletal muscle fibers will contract without extracellular
calcium.
During contraction of a cardiac muscle cell, the long protein myofilaments oriented along the length of the cell slide over each other in what is known as the sliding filament theory. There are two kinds of myofilaments, thick filaments composed of the protein myosin, and thin filaments composed of the proteins actin, troponin and tropomyosin. As the thick and thin filaments slide past each other the cell becomes shorter and fatter. In a mechanism known as cross-bridge cycling,
calcium ions bind to the protein troponin, which along with tropomyosin
then uncover key binding sites on actin. Myosin, in the thick
filament, can then bind to actin, pulling the thick filaments along the
thin filaments. When the concentration of calcium within the cell
falls, troponin and tropomyosin once again cover the binding sites on
actin, causing the cell to relax.
Regeneration
Dog cardiac muscle (400X)
It was commonly believed that cardiac muscle cells could not be
regenerated. However, this was contradicted by a report published in
2009. Olaf Bergmann and his colleagues at the Karolinska Institute in Stockholm
tested samples of heart muscle from people born before 1955 who had
very little cardiac muscle around their heart, many showing with
disabilities from this abnormality. By using DNA samples from many
hearts, the researchers estimated that a 4-year-old renews about 20% of
heart muscle cells per year, and about 69 percent of the heart muscle
cells of a 50-year-old were generated after he or she was born.
One way that cardiomyocyte regeneration occurs is through the
division of pre-existing cardiomyocytes during the normal aging process.
In the 2000s, the discovery of adult endogenous cardiac stem
cells was reported, and studies were published that claimed that various
stem cell lineages, including bone marrow stem cells were able to differentiate into cardiomyocytes, and could be used to treat heart failure.
However, other teams were unable to replicate these findings, and many of the original studies were later retracted for scientific fraud.
Differences between atria and ventricles
The swirling musculature of the heart ensures effective pumping of blood.
Cardiac muscle forms both the atria and the ventricles of the heart.
Although this muscle tissue is very similar between cardiac chambers,
some differences exist. The myocardium found in the ventricles is thick
to allow forceful contractions, while the myocardium in the atria is
much thinner. The individual myocytes that make up the myocardium also
differ between cardiac chambers. Ventricular cardiomyocytes are longer
and wider, with a denser T-tubule
network. Although the fundamental mechanisms of calcium handling are
similar between ventricular and atrial cardiomyocytes, the calcium
transient is smaller and decays more rapidly in atrial myocytes, with a
corresponding increase in calcium buffering capacity.
The complement of ion channels differs between chambers, leading to
longer action potential durations and effective refractory periods in
the ventricles. Certain ion currents such as IK(UR) are highly specific to atrial cardiomyocytes, making them a potential target for treatments for atrial fibrillation.
Heart muscle can also become damaged despite a normal blood supply. The heart muscle may become inflamed in a condition called myocarditis, most commonly caused by a viral infection but sometimes caused by the body's own immune system. Heart muscle can also be damaged by drugs such as alcohol, long standing high blood pressure or hypertension, or persistent abnormal heart racing.
Many of these conditions, if severe enough, can damage the heart so much
that the pumping function of the heart is reduced. If the heart is no
longer able to pump enough blood to meet the body's needs, this is
described as heart failure.
Significant damage to cardiac muscle cells is referred to as myocytolysis which is considered a type of cellular necrosis defined as either coagulative or colliquative.
The laws describing the behaviour of gases under fixed pressure, volume and absolutetemperature conditions are called Gas Laws.
The basic gas laws were discovered by the end of the 18th century when
scientists found out that relationships between pressure, volume and
temperature of a sample of gas could be obtained which would hold to
approximation for all gases. These macroscopic gas laws were found to be
consistent with atomic and kinetic theory.
History
Following the invention of the Torricelli mercury barometer in mid
17th century, the pressure-volume gas law was soon revealed by Robert Boyle
while keeping temperature constant. Marriott, however, did notice small
temperature dependence. It took another century and a half to develop
thermometry and recognise the absolute zero temperature scale before the
discovery of temperature-dependent gas laws.
In 1662, Robert Boyle systematically studied the relationship between
the volume and pressure of a fixed amount of gas at a constant
temperature. He observed that the volume of a given mass of a gas is
inversely proportional to its pressure at a constant temperature.
Boyle's law, published in 1662, states that, at a constant temperature,
the product of the pressure and volume of a given mass of an ideal gas in a closed system is always constant. It can be verified experimentally using a pressure gauge and a variable volume container. It can also be derived from the kinetic theory of gases: if a container, with a fixed number of molecules
inside, is reduced in volume, more molecules will strike a given area
of the sides of the container per unit time, causing a greater pressure.
Statement
Boyle's law states that:
The volume of a given mass of a dry gas is inversely related to its pressure when its temperature is kept constant.
The concept can be represented with these formulae:
, meaning "Volume is inversely proportional to Pressure", or
, meaning "Pressure is inversely proportional to Volume", or
, or
where P is the pressure, V is the volume of a gas, and k1 is the constant in this equation (and is not the same as the proportionality constants in the other equations).
Charles' law, or the law of volumes, was founded in 1787 by Jacques Charles. It states that, for a given mass of an ideal gas at constant pressure, the volume is directly proportional to its absolute temperature, assuming in a closed system.
The statement of Charles' law is as follows:
the volume (V) of a given mass of a gas, at constant pressure (P), is directly proportional to its temperature (T).
Statement
Charles' law states that:
The volume of a given fixed mass of a dry gas is directly proportional to its absolute temperature at a constant pressure.
Therefore,
, or
, or
,
where "V" is the volume of a gas, "T" is the absolute temperature and k2 is a proportionality constant (which is not the same as the proportionality constants in the other equations in this article).
Gay-Lussac's law, Amontons' law or the pressure law was founded by Joseph Louis Gay-Lussac in 1808.
Statement
Gay-Lussac's law states that:
The pressure exerted by a given mass and
constant volume of an ideal gas on the sides of its container is
directly proportional to its absolute temperature.
Therefore,
, or
, or
,
where P is the pressure, T is the absolute temperature, and k is another proportionality constant.
Avogadro's law, Avogadro's hypothesis, Avogadro's principle or Avogadro-Ampère's hypothesis is an experimental gas law which was hypothesized by Amedeo Avogadro in 1811. It related the volume of a gas to the amount of substance of gas present.
Statement
Avogadro's law states that:
The volume occupied by an ideal gas at a
constant temperature is directly proportional to the number of molecules
of the gas present in the container.
This statement gives rise to the molar volume of a gas, which at STP (273.15 K, 1 atm) is about 22.4 L. The relation is given by:
, or
where n is equal to the number of molecules of gas (or the number of moles of gas).
The Combined gas law
or General Gas Equation is obtained by combining Boyle's Law, Charles's
law, and Gay-Lussac's Law. It shows the relationship between the
pressure, volume, and temperature for a fixed mass of gas:
where P is the pressure, V is volume, n is the number of moles, R is the universal gas constant and T is the absolute temperature.
The proportionality constant, now named R, is the universal gas constant with a value of 8.3144598 (kPa∙L)/(mol∙K).
An equivalent formulation of this law is:
where P is the pressure, V is the volume, N is the number of gas molecules, kB is the Boltzmann constant (1.381×10−23J·K−1 in SI units) and T is the absolute temperature.
These equations are exact only for an ideal gas, which neglects various intermolecular effects (see real gas). However, the ideal gas law is a good approximation for most gases under moderate pressure and temperature.
This law has the following important consequences:
If temperature and pressure are kept constant, then the volume
of the gas is directly proportional to the number of molecules of gas.
If the temperature and volume remain constant, then the pressure of
the gas changes is directly proportional to the number of molecules of
gas present.
If the number of gas molecules and the temperature remain constant, then the pressure is inversely proportional to the volume.
If the temperature changes and the number of gas molecules are kept
constant, then either pressure or volume (or both) will change in direct
proportion to the temperature.
This law states that the rate at which gas molecules diffuse
is inversely proportional to the square root of the gas density at a
constant temperature. Combined with Avogadro's law (i.e. since equal
volumes have an equal number of molecules) this is the same as being
inversely proportional to the root of the molecular weight.
This law states that the pressure of a mixture of gases simply is the sum of the partial pressures of the individual components. Dalton's law is as follows:
and all component gases and the mixture are at the same temperature and volume
where Ptotal is the total pressure of the gas mixture
Pi is the partial pressure or pressure of the component gas at the given volume and temperature.
This law states that the volume of a mixture of gases (or the volume
of the container) simply is the sum of the partial volumes of the
individual components. Amagat's law is as follows:
and all component gases and the mixture are at the same temperature and pressure
where Vtotal is the total volume of the gas mixture or the volume of the container,
Vi is the partial volume, or volume of the component gas at the given pressure and temperature.
states that At constant temperature, the amount of a given gas
dissolved in a given type and volume of liquid is directly proportional
to the partial pressure of that gas in equilibrium with that liquid.