Effects of cognitive rehabilitation therapy, assessed using fMRI.
Cognitive rehabilitation refers to a wide range of evidence-based interventions
designed to improve cognitive functioning in brain-injured or otherwise
cognitively impaired individuals to restore normal functioning, or to
compensate for cognitive deficits. It entails an individualized program of specific skills training and practice plus metacognitive strategies.
Metacognitive strategies include helping the patient increase
self-awareness regarding problem-solving skills by learning how to
monitor the effectiveness of these skills and self-correct when
necessary.
Cognitive rehabilitation therapy (offered by a trained therapist)
is a subset of Cognitive Rehabilitation (community-based
rehabilitation, often in traumatic brain injury; provided by
rehabilitation professionals) and has been shown to be effective for
individuals who had a stroke in the left or right hemisphere. or brain trauma. A computer-assisted type of cognitive rehabilitation therapy called cognitive remediation therapy has been used to treat schizophrenia, ADHD, and major depressive disorder.
It may also be recommended for traumatic brain injury, the primary population for which it was developed in the university medical and rehabilitation communities, such as that sustained by U.S. Representative Gabby Giffords, according to Dr. Gregory J. O'Shanick of the Brain Injury Association of America. Her new doctor has confirmed that it will be part of her rehabilitation.
Cognitive rehabilitation may be part of a comprehensive community
services program and integrated into residential services, such as supported living, supported employment, family support, professional education, home health (as personal assistance),recreation, or education programs in the community.
Cognitive rehabilitation for spatial neglect following stroke
The current body of evidence is uncertain on the efficacy of
cognitive rehabilitation for reducing the disabling effects of neglect
and increasing independence remains unproven. However, there is limited evidence that cognitive rehabilitation may have an immediate beneficial effect on tests of neglect. Overall, no rehabilitation approach can be supported by evidence for spatial neglect.
Assessments
According to the standard text by Sohlberg and Mateer:
Individuals
and families respond differently to different interventions, in
different ways, at different times after injury. Premorbid functioning,
personality, social support, and environmental demands are but a few of
the factors that can profoundly influence outcome. In this variable
response to treatment, cognitive rehabilitation is no different from
treatment for cancer, diabetes, heart disease, Parkinson's disease,
spinal cord injury, psychiatric disorders, or any other injury or
disease process for which variable response to different treatments is
the norm.
Nevertheless, many different statistical analyses of the benefits of
this therapy have been carried out. One study made in 2002 analyzed 47
treatment comparisons and reported "a differential benefit in favor of
cognitive rehabilitation in 37 of 47 (78.7%) comparisons, with no
comparison demonstrating a benefit in favor of the alternative treatment
condition."
An internal study conducted by the Tricare Management Agency in 2009 is
cited by the US Department of Defense as its reason for refusing to pay
for this therapy for veterans who have had traumatic brain injury.
According to Tricare, "There is insufficient, evidence-based research
available to conclude that cognitive rehabilitation therapy is
beneficial in treating traumatic brain injury."
The ECRI Institute, whose report serves as the basis for this decision
by the Department of Defense, has summed up their own findings this way:
In
our report, we carried out several meta-analyses using data from 18
randomized controlled trials. Based on data from these studies, we were
able to conclude the following:
Adults with moderate to severe traumatic brain injury who
receive social skills training perform significantly better on measures
of social communication than patients who receive no treatment.
Adults with traumatic brain injury who receive comprehensive
cognitive rehabilitation therapy report significant improvement on
measures of quality of life compared to patients who receive a less
intense form of therapy.
The strength of the evidence supporting our conclusions was low due to
the small number of studies that addressed the outcomes of interest.
Further, the evidence was too weak to draw any definitive conclusions
about the effectiveness of cognitive rehabilitation therapy for treating
deficits related to the following cognitive areas: attention, memory,
visuospacial skills, and executive function. The following factors
contributed to the weakness of the evidence: differences in the outcomes
assessed in the studies, differences in the types of cognitive
rehabilitation therapy methods/strategies employed across studies,
differences in the control conditions, and/or insufficient number of
studies addressing an outcome.
Citing this 2009 assessment, US Department of Defense,
one of the federal agencies not responsible for health care decisions
in the US, has declared that cognitive rehabilitation therapy is
scientifically unproved and should refer their concerns to the US
Department of Health and Human Services, US Budget and Management,
and/or the Government Accountability Office (GAO). As a result, it
refuses to cover the cost of cognitive rehabilitation for brain-injured
veterans.
Cost-benefit and cost-effectiveness studies, together with an analysis
of personnel and veterans' services for new our emerging groups in head
and brain injuries, are recommended.
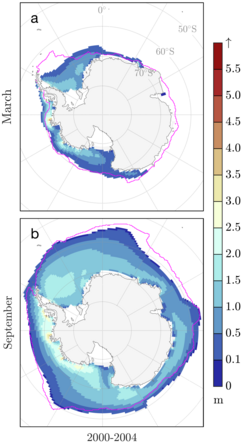
CICE (/saɪs/) is a computer model that simulates the growth, melt and movement of sea ice. It has been integrated into many coupled climate system models as well as global ocean and weather forecasting models and is often used as a tool in Arctic and Southern Ocean research. CICE development began in the mid-1990s by the United States Department of Energy
(DOE), and it is currently maintained and developed by a group of
institutions in North America and Europe known as the CICE Consortium. Its widespread use in Earth system science in part owes to the importance of sea ice in determining Earth's planetary albedo, the strength of the global thermohaline circulation in the world's oceans, and in providing surface boundary conditions for atmospheric circulation models, since sea ice occupies a significant proportion (4-6%) of Earth's surface. CICE is a type of cryospheric model.
where is the mass per unit area of saline ice on the sea surface, is the drift velocity of the ice, is the Coriolis parameter, is the upward unit vector normal to the sea surface, and are the wind and water stress on the ice, respectively, is acceleration due to gravity, is sea surface height and is internal ice the two-dimensional stress tensor within the ice.
Each of the terms require information about the ice thickness,
roughness, and concentration, as well as the state of the atmospheric
and oceanic boundary layers. Ice mass per unit area is determined using the second keystone equation in CICE, which describes evolution of the sea ice thickness distribution for different thicknesses spread of the area for which sea ice velocity is calculated above:
where is the change in the thickness distribution due to thermodynamic growth and melt, is redistribution function due to sea ice mechanics and is associated with internal ice stress , and describes advection of sea ice in a Lagrangian reference frame. From this, ice mass is given by:
Schematic demonstrating placement of Icepack, in which the thickness distribution
is represented (blue), within the MPAS dycore (green) that solves for
momentum evolution and horizontal sea ice advection on the E3SM
unstructured grid (arrows)
CICE version 6 is coded in FORTRAN90. It is organized into a dynamical core (dycore) and a separate column physics package called Icepack, which is maintained as a CICE submodule on GitHub. The momentum equation and thickness advection described above are time-stepped on a quadrilateralArakawa B-grid
within the dynamical core, while Icepack solves diagnostic and
prognostic equations necessary for calculating radiation physics,
hydrology, thermodynamics, and vertical biogeochemistry, including terms
necessary to calculate , , , , and
defined above. CICE can be run independently, as in the first figure on
this page, but is frequently coupled with earth systems models through
an external flux coupler, such as the CESM Flux Coupler from NCAR
for which results are shown in the second figure for the CESM Large
Ensemble. The column physics were separated into Icepack for the version
6 release to permit insertion into earth system models that use their
own sea ice dynamical core, including the new DOE Energy Exascale Earth
System Model (E3SM), which uses an unstructured grid in the sea ice component of the Model for Prediction Across Scales (MPAS), as demonstrated in the final figure.
A stressor is a chemical or biological agent, environmental condition, external stimulus or an event seen as causing stress to an organism.
Psychologically speaking, a stressor can be events or environments that
individuals might consider demanding, challenging, and/or threatening
individual safety.
Events or objects that may trigger a stress response may include:
Stressors can cause physical, chemical and mental responses
internally. Physical stressors produce mechanical stresses on skin,
bones, ligaments, tendons, muscles and nerves that cause tissue
deformation and (in extreme cases) tissue failure. Chemical stresses
also produce biomechanical responses associated with metabolism
and tissue repair. Physical stressors may produce pain and impair work
performance. Chronic pain and impairment requiring medical attention
may result from extreme physical stressors or if there is not sufficient
recovery time between successive exposures.
Stressors may also affect mental function and performance. Mental and
social stressors may affect behavior and how individuals respond to
physical and chemical stressors.
Social and environmental stressors and the events associated with
them can range from minor to traumatic. Traumatic events involve very
debilitating stressors, and oftentimes these stressors are
uncontrollable. Traumatic events can deplete an individual's coping resources to an extent where the individual may develop acute stress disorder or even post-traumatic stress disorder. People who have been abused, victimized, or terrorized are often more susceptible to stress disorders.
Most stressor-stress relationships can be evaluated and determined -
either by the individual or by a psychologist. Therapeutic measures are
often taken to help replenish and rebuild the individual's coping
resources while simultaneously aiding the individual in dealing with
current stress.
Psychological stressors
Stressors
occur when an individual is unable to cope with the demands of their
environment (such as crippling debt with no clear path to resolving it).
Generally, stressors take many forms, such as: traumatic events, life
demands, sudden medical emergencies, and daily inconveniences, to name a
few. There are also a variety of characteristics that a stressor may
possess (different durations, intensity, predictability, and
controllability).
Measuring psychological stress
Due
to the wide impact and the far-reaching consequences of psychological
stressors (especially their profound effects on mental well-being), it
is particularly important to devise tools to measure such stressors. Two
common psychological stress tests include the Perceived Stress Scale (PSS) devised by American psychologist Sheldon Cohen, and the Social Readjustment Rating Scale (SRRS) or the Holmes-Rahe Stress Scale. While the PSS is a traditional Likert scale, the SRRS assigns specific predefined numerical values to stressors.
Biological responses to stressors
Traumatic
events or any type of shock to the body can cause an acute stress
response disorder (ASD). The extent to which one experiences ASD depends
on the extent of the shock. If the shock was pushed past a certain
extreme after a particular period in time ASD can develop into what is
commonly known as Post-traumatic stress disorder (PTSD).
There are two ways that the body responds biologically in order to
reduce the amount of stress an individual is experiencing. One thing
that the body does to combat stressors is to create stress hormones,
which in turn create energy reservoirs that are there in case a
stressful event were to occur. The second way our biological components
respond is through an individual's cells. Depending on the situation our
cells obtain more energy in order to combat any negative stressor and
any other activity those cells are involved in seize.
When
individuals are informed about events before they occur, the magnitude
of the stressor is less than when compared to individuals who were not
informed of the stressor.
For example, an individual would prefer to know when they have a
deadline ahead of time in order to prepare for it in advance, rather
than find out about the deadline the day of. In knowing that there is a
deadline ahead of time, the intensity of the stressor is smaller for the
individual, as opposed to the magnitude of intensity for the other
unfortunate individual who found out about the deadline the day of. When
this was tested, psychologists found that when given the choice,
individuals had a preference for the predictable stressors, rather than
the unpredictable stressors. The pathologies caused by the lack of predictability are experienced by some individuals working in fields of emergency medicine, military defense, disaster response and others.
Additionally, the degree to which the stressor can be controlled plays a variable in how the individual perceives stress.
Research has found that if an individual is able to take some control
over the stressor, then the level of stress will be decreased. During
this study, it was found that the individuals become increasingly
anxious and distressed if they were unable to control their environment.
As an example, imagine an individual who detests baths in the Middle
Ages, taking a bath. If the individual was forced to take the bath with
no control over the temperature of the bath (one of the variables), then
their anxiety and stress levels would be higher than if the individual
was given some control over the environment (such as being able to
control the temperature of the water).
Based on these two principles (predictability and control), there
are two hypotheses that attempt to account for these preferences; the
preparatory response hypothesis and safety hypothesis attempt to
accommodate these preferences.
Preparatory response hypothesis
The
idea behind this hypothesis is that an organism can better prepare for
an event if they are informed beforehand, as this allows them to prepare
for it (biologically). In biologically preparing for this event beforehand, the individual is able to better decrease the event's aversiveness.
In knowing when a potential stressor will occur (such as an exam), the
individual could, in theory, prepare for it in advance, thus decreasing
the stress that may result from that event.
Safety hypothesis
In
this hypothesis, there are two time periods, one in which is deemed
safe (where there is no stressor), and one which is deemed unsafe (in
which the stressor is present).
This is similar to procrastination and cramming; during the safe
intervals (weeks before an exam) the individual is relaxed and not
anxious, and during the unsafe intervals (the day or night before the
exam) the individual most likely experiences anxiety.
Diagram of a class-C stellar engine (to scale) built around a Sun-like star. It consists of a partial Dyson swarm composed of 5 Dyson rings of solar collectors (the class-B component), and a large statite Shkadov thruster (the class-A component). Perspective is from below the system's ecliptic at a distance of ~2.8 AU. The system's direction of acceleration is on a vector
from the center of the star through the center of the Shkadov thruster,
which is hovering over the star's north pole (with regards to the
ecliptic), at a distance of 1 AU.
Stellar engines are a class of hypothetical megastructures which use the resources of a star to generate available work (also called exergy).
For instance, they can use the energy of the star to produce
mechanical, electrical or chemical work or they can use the impulse of
the light emitted by the star to produce thrust, able to control the
motion of a star system. The concept has been introduced by Bădescu and Cathcart. The variants which produce thrust may accelerate a star and anything orbiting it in a given direction. The creation of such a system would make its builders a type-II civilization on the Kardashev scale.
Classes
Three classes of stellar engines have been defined.
Class A (Shkadov thruster)
One of the simplest examples of a stellar engine is the Shkadov thruster (named after Dr. Leonid Shkadov, who first proposed it), or a class-A stellar engine. Such an engine is a stellar propulsion system, consisting of an enormous mirror/light sail—actually a massive type of solar statite large enough to classify as a megastructure—which would balance gravitational attraction towards and radiation pressure away from the star. Since the radiation pressure of the star would now be asymmetrical, i.e. more radiation being emitted in one direction as compared to another, the "excess" radiation pressure acts as net thrust, accelerating the star in the direction of the hovering statite. Such thrust and acceleration would be very slight, but such a system could be stable for millennia. Any planetary system attached to the star would be "dragged" along by its parent star. For a star such as the Sun, with luminosity 3.85×1026W and mass 1.99×1030kg, the total thrust produced by reflecting half of the solar output would be 1.28×1018N.
After a period of one million years this would yield an imparted speed
of 20 m/s, with a displacement from the original position of 0.03 light-years.
After one billion years, the speed would be 20 km/s and the
displacement 34,000 light-years, a little over a third of the estimated
width of the Milky Way galaxy.
Class B
A class-B
stellar engine consists of two concentric spheres around a star. The
inner sphere (which may be assimilated with a Dyson shell) receives
energy from the star and becomes hotter than the outer sphere. The
difference of temperature between the two spheres drives thermal engines
able to provide mechanical work.
Unlike the Shkadov thruster, a class-B stellar engine is not propulsive.
Class C
A class-C stellar engine, such as the Badescu–Cathcart engine,
combines the two other classes, employing both the propulsive aspects
of the Shkadov thruster and the energy generating aspects of a class-B
engine. A higher temperature Dyson shell
partially covered by a mirror combined with an outer sphere at a lower
temperature would be one incarnation of such a system. The non-spherical
mirror ensures conversion of light impulse into effective thrust
(like a class-A stellar engine) while the difference of temperature may
be used to convert star energy into mechanical work (like a class-B
stellar engine). Notice that such system suffers from the same
stabilization problems as a non-propulsive shell, as would be a Dyson swarm with a large statite mirror (see image above). A Dyson bubble
variant is already a Shkadov thruster (provided that the arrangement of
statite components is asymmetrical); adding energy extraction
capability to the components seems an almost trivial extension.
Caplan thruster
Astronomer Matthew E. Caplan of Illinois State University
has proposed a type of stellar engine that uses concentrated stellar
energy (repurposing the mirror statites from class A) to excite certain
regions of the outer surface of the star and create beams of solar wind for collection by a multi-Bussard ramjet assembly. The ramjets would produce directed plasma to stabilize its orbit and jets of oxygen-14
to push the star. Using rudimentary calculations that assume maximum
efficiency, Caplan estimates that the Bussard engine would use 1012 kg of solar material per second to produce a maximum acceleration of 10−9 m/s2, yielding a velocity of 200 km/s after 5 million years and a distance of 10 parsecs
over 1 million years. While theoretically the Bussard engine would work
for 100 million years, given the mass loss rate of the Sun, Caplan
deems 10 million years to be sufficient for a stellar collision
avoidance. His proposal was commissioned by the German educational YouTube channel Kurzgesagt.
Svoronos Star Tug
Alexander
A. Svoronos of Yale University proposed the 'Star Tug', a concept that
combines aspects of the Shkadov thruster and Caplan engine to produce an
even more powerful and efficient mechanism for controlling a star's
movement. Essentially, it replaces the giant parabolic mirror of the
Shkadov thruster with an engine powered by mass lifted from the star,
similar to the Caplan engine. However, instead of pushing a star from
behind with a beam of thrust, as the Caplan engine does, it pulls the
star from the front via its gravitational link to it, same as the
Shkadov thruster. As a result, it only needs to produce a single beam of
thrust (toward but narrowly missing the star), whereas the Caplan
engine must produce two beams of thrust (one to push the star from
behind and negate the force of gravity between the engine and the star,
and one to propel the system as a whole forward). The result is that the
Svoronos Star Tug is a much more efficient engine capable of
significantly higher accelerations and max velocities. The Svoronos Star
Tug can, in principle (assuming perfect efficiency), accelerate the Sun
to ~27% the speed of light (after burning enough of the Sun's mass to
transition it to a brown dwarf).
Bile acids are steroidacids found predominantly in the bile of mammals and other vertebrates. Diverse bile acids are synthesized in the liver. Bile acids are conjugated with taurine or glycine residues to give anions called bile salts.
Bile acids comprise about 80% of the organic compounds in bile (others are phospholipids and cholesterol). An increased secretion of bile acids produces an increase in bile flow. Bile acids facilitate digestion of dietary fats and oils. They serve as micelle-forming surfactants, which encapsulate nutrients, facilitating their absorption. These micelles are suspended in the chyme before further processing. Bile acids also have hormonal actions throughout the body, particularly through the farnesoid X receptor and GPBAR1 (also known as TGR5).
Bile acid synthesis is the only manner in which humans or other
mammals may excrete excess cholesterol, as the parent compound of all
bile acids is cholesterol.
Structure of cholic acid showing relationship to other bile acids
Production
Primary bile acids
Bile acid synthesis occurs in liver cells, which synthesize primary bile acids (cholic acid and chenodeoxycholic acid in humans) via cytochrome P450-mediated oxidation of cholesterol
in a multi-step process. Approximately 600 mg of bile salts are
synthesized daily to replace bile acids lost in the feces, although, as
described below, much larger amounts are secreted, reabsorbed in the gut
and recycled.
Prior to secreting any of the bile acids (primary or secondary, see below), liver cells conjugate them with either glycine or taurine, to form a total of 8 possible conjugated bile acids. These conjugated bile acids are often referred to as bile salts. The pKa of the unconjugated bile acids are between 5 and 6.5,
and the pH of the duodenum ranges between 3 and 5, so when unconjugated
bile acids are in the duodenum, they are almost always protonated (HA
form), which makes them relatively insoluble in water. Conjugating bile
acids with amino acids lowers the pKa of the bile-acid/amino-acid
conjugate to between 1 and 4. Thus conjugated bile acids are almost
always in their deprotonated (A-) form in the duodenum, which makes them
much more water-soluble and much more able to fulfil their physiologic
function of emulsifying fats.
Secondary bile acids
Once
secreted into the lumen of the intestine, bile salts are modified by
gut bacteria. They are partially dehydroxylated. Their glycine and
taurine groups are removed to give the secondary bile acids, deoxycholic acid and lithocholic acid.
Cholic acid is converted into deoxycholic acid and chenodeoxycholic
acid into lithocholic acid. All four of these bile acids are recycled,
in a process known as enterohepatic circulation.
Functions
Lipid digestion
As molecules with hydrophobic and hydrophilic regions, conjugated bile salts sit at the lipid/water interface and, above the right concentration, form micelles.
The added solubility of conjugated bile salts aids in their function by
preventing passive re-absorption in the small intestine. As a result,
the concentration of bile acids/salts in the small intestine is high
enough to form micelles and solubilize lipids. "Critical micellar
concentration" refers to both an intrinsic property of the bile acid
itself and amount of bile acid necessary to function in the spontaneous
and dynamic formation of micelles. Bile acid-containing micelles aid lipases to digest lipids and bring them near the intestinal brush border membrane, which results in fat absorption.
Synthesis of bile acids is a major route of cholesterol
metabolism in most species other than humans. The body produces about
800 mg of cholesterol per day and about half of that is used for bile
acid synthesis producing 400–600 mg daily. Human adults secrete between
12 and 18 g of bile acids into the intestine each day, mostly after
meals. The bile acid pool size is between 4–6 g, which means that bile
acids are recycled several times each day. About 95% of bile acids are
reabsorbed by active transport in the ileum and recycled back to the liver for further secretion into the biliary system and gallbladder. This enterohepatic circulation of bile acids allows a low rate of synthesis, only about 0.3 g/day, but with large amounts being secreted into the intestine.
Bile acids have other functions, including eliminating
cholesterol from the body, driving the flow of bile to eliminate certain
catabolites (including bilirubin),
emulsifying fat-soluble vitamins to enable their absorption, and aiding
in motility and the reduction of the bacteria flora found in the small
intestine and biliary tract.
Cell signalling
Bile acids have metabolic actions in the body resembling those of hormones, acting through two specific receptors, the farnesoid X receptor and G protein-coupled bile acid receptor/TGR5. They bind less specifically to some other receptors and have been reported to regulate the activity of certain enzymes and ion channels and the synthesis of diverse substances including endogenous fatty acid ethanolamides.
Bile salts constitute a large family of molecules, composed of a
steroid structure with four rings, a five- or eight-carbon side-chain
terminating in a carboxylic acid, and several hydroxyl groups, the
number and orientation of which is different among the specific bile
salts.
The four rings are labeled A, B, C, and D, from the farthest to the
closest to the side chain with the carboxyl group. The D-ring is smaller
by one carbon than the other three. The structure is commonly drawn
with A at the left and D at the right. The hydroxyl groups can be in
either of two configurations: either up (or out), termed beta (β; often
drawn by convention as a solid line), or down, termed alpha (α;
displayed as a dashed line). All bile acids have a 3-hydroxyl group,
derived from the parent molecule, cholesterol, in which the 3-hydroxyl
is beta.
IUPAC recommended ring lettering (left) and atom numbering (right) of the steroid skeleton. The four rings A-D form a sterane core.
The initial step in the classical pathway of hepatic synthesis of
bile acids is the enzymatic addition of a 7α hydroxyl group by cholesterol 7α-hydroxylase (CYP7A1) forming 7α-hydroxycholesterol. This is then metabolised to 7α-hydroxy-4-cholesten-3-one. There are multiple steps in bile acid synthesis requiring 14 enzymes in all.
These result in the junction between the first two steroid rings (A and
B) being altered, making the molecule bent; in this process, the
3-hydroxyl is converted to the α orientation. The simplest 24-carbon
bile acid has two hydroxyl groups at positions 3α and 7α. This is
3α,7α-dihydroxy-5β-cholan-24-oic acid, or, as more usually known, chenodeoxycholic acid. This bile acid was first isolated from the domestic goose,
from which the "cheno" portion of the name was derived (Greek: χήν =
goose). The 5β in the name denotes the orientation of the junction
between rings A and B of the steroid nucleus (in this case, they are
bent). The term "cholan" denotes a particular steroid structure of 24
carbons, and the "24-oic acid" indicates that the carboxylic acid is
found at position 24, at the end of the side-chain. Chenodeoxycholic
acid is made by many species, and is the prototypic functional bile
acid.
An alternative (acidic) pathway of bile acid synthesis is initiated by mitochondrial sterol 27-hydroxylase (CYP27A1),
expressed in liver, and also in macrophages and other tissues. CYP27A1
contributes significantly to total bile acid synthesis by catalyzing
sterol side chain oxidation, after which cleavage of a three-carbon unit
in the peroxisomes
leads to formation of a C24 bile acid. Minor pathways initiated by
25-hydroxylase in the liver and 24-hydroxylase in the brain also may
contribute to bile acid synthesis. 7α-hydroxylase (CYP7B1) generates oxysterols, which may be further converted in the liver to CDCA.
Cholic acid,
3α,7α,12α-trihydroxy-5β-cholan-24-oic acid, the most abundant bile acid
in humans and many other species, was discovered before
chenodeoxycholic acid. It is a tri-hydroxy-bile acid with 3 hydroxyl
groups (3α, 7α and 12α). In its synthesis in the liver, 12α
hydroxylation is performed by the additional action of CYP8B1.
As this had already been described, the discovery of chenodeoxycholic
acid (with 2 hydroxyl groups) made this new bile acid a "deoxycholic
acid" in that it had one fewer hydroxyl group than cholic acid.
Deoxycholic acid
is formed from cholic acid by 7-dehydroxylation, resulting in 2
hydroxyl groups (3α and 12α). This process with chenodeoxycholic acid
results in a bile acid with only a 3α hydroxyl group, termed lithocholic acid (litho = stone) having been identified first in a gallstone from a calf. It is poorly water-soluble and rather toxic to cells.
Different vertebrate families have evolved to use modifications
of most positions on the steroid nucleus and side-chain of the bile acid
structure. To avoid the problems associated with the production of
lithocholic acid, most species add a third hydroxyl group to
chenodeoxycholic acid. The subsequent removal of the 7α hydroxyl group
by intestinal bacteria will then result in a less toxic but
still-functional dihydroxy bile acid. Over the course of vertebrate
evolution, a number of positions have been chosen for placement of the
third hydroxyl group. Initially, the 16α position was favored, in
particular in birds. Later, this position was superseded in a large
number of species selecting the 12α position. Primates (including
humans) utilize 12α for their third hydroxyl group position, producing
cholic acid. In mice and other rodents, 6β hydroxylation forms muricholic acids (α or β depending on the 7 hydroxyl position). Pigs have 6α hydroxylation in hyocholic acid (3α,6α,7α-trihydroxy-5β-cholanoic acid), and other species have a hydroxyl group on position 23 of the side-chain.
Many other bile acids have been described, often in small
amounts, resulting from bacterial enzymatic or other modifications. The
"iso-" epimers have the 3-hydroxyl group in the β position. The "allo-"
epimers have the 5α configuration, which changes the relative position
of the A and B rings.
Ursodeoxycholic acid was first isolated from bear bile,
which has been used medicinally for centuries. Its structure resembles
chenodeoxycholic acid but with the 7-hydroxyl group in the β position.
Obeticholic acid,
6α-ethyl-chenodeoxycholic acid, is a semi-synthetic bile acid with
greater activity as an FXR agonist, which has been developed as a
pharmaceutical agent in certain liver diseases.
Hormonal actions
Bile
acids also act as steroid hormones, secreted from the liver, absorbed
from the intestine and having various direct metabolic actions in the
body through the nuclear receptor Farnesoid X receptor (FXR), also known by its gene name NR1H4. Another bile acid receptor is the cell membrane receptor known as G protein-coupled bile acid receptor 1 or TGR5.
Many of their functions as signaling molecules in the liver and the
intestines are by activating FXR, whereas TGR5 may be involved in
metabolic, endocrine and neurological functions.
Regulation of synthesis
As surfactants or detergents, bile acids are potentially toxic to cells, and so their concentrations
are tightly regulated. Activation of FXR in the liver inhibits
synthesis of bile acids, and is one mechanism of feedback control when
bile acid levels are too high. Secondly, FXR activation by bile acids
during absorption in the intestine increases transcription and synthesis
of FGF19, which then inhibits bile acid synthesis in the liver.
Bile
acids bind to some other proteins in addition to their hormone
receptors (FXR and TGR5) and their transporters. Among these protein
targets, the enzyme N-acyl phosphatidylethanolamine-specific phospholipase D (NAPE-PLD) generates bioactive lipid amides (e.g. the endogenous cannabinoidanandamide) that play important roles in several physiological pathways including stress and pain responses, appetite, and lifespan. NAPE-PLD orchestrates a direct cross-talk between lipid amide signals and bile acid physiology.
Clinical significance
Hyperlipidemia
As
bile acids are made from endogenous cholesterol, disruption of the
enterohepatic circulation of bile acids will lower cholesterol. Bile acid sequestrants
bind bile acids in the gut, preventing reabsorption. In so doing, more
endogenous cholesterol is shunted into the production of bile acids,
thereby lowering cholesterol levels. The sequestered bile acids are then
excreted in the feces.
The relationship of bile acids to cholesterol saturation in bile and
cholesterol precipitation to produce gallstones has been studied
extensively. Gallstones may result from increased saturation of
cholesterol or bilirubin,
or from bile stasis. Lower concentrations of bile acids or
phospholipids in bile reduce cholesterol solubility and lead to
microcrystal formation. Oral therapy with chenodeoxycholic acid and/or
ursodeoxycholic acid has been used to dissolve cholesterol gallstones.
Stones may recur when treatment is stopped. Bile acid therapy may be of
value to prevent stones in certain circumstances such as following bariatric surgery.
Bile acids may have some importance in the development of colorectal cancer. Deoxycholic acid (DCA) is increased in the colonic contents of humans in response to a high fat diet. In populations with a high incidence of colorectal cancer, fecal concentrations of bile acids are higher,
and this association suggests that increased colonic exposure to bile
acids could play a role in the development of cancer. In one particular
comparison, the fecal DCA concentrations in Native Africans in South
Africa (who eat a low fat diet) compared to African Americans (who eat a
higher fat diet) was 7.30 vs. 37.51 nmol/g wet weight stool. Native Africans in South Africa have a low incidence rate of colon cancer of less than 1:100,000, compared to the high incidence rate for male African Americans of 72:100,000.
Experimental studies also suggest mechanisms for bile acids in
colon cancer. Exposure of colonic cells to high DCA concentrations
increase formation of reactive oxygen species, causing oxidative stress, and also increase DNA damage. Mice fed a diet with added DCA mimicking colonic DCA levels in humans on a high fat diet developed colonic neoplasia, including adenomas and adenocarcinomas (cancers), unlike mice fed a control diet producing one-tenth the level of colonic DCA who had no colonic neoplasia.
The effects of ursodeoxycholic acid (UDCA) in modifying the risk
of colorectal cancer has been looked at in several studies, particularly
in primary sclerosing cholangitis and inflammatory bowel disease, with varying results partly related to dosage. Genetic variation in the key bile acid synthesis enzyme, CYP7A1, influenced the effectiveness of UDCA in colorectal adenoma prevention in a large trial.
Dermatology
Bile acids may be used in subcutaneous injections to remove unwanted fat (see Mesotherapy). Deoxycholic acid as an injectable has received FDA approval to dissolve submental fat.
Phase III trials showed significant responses although many subjects
had mild adverse reactions of bruising, swelling, pain, numbness,
erythema, and firmness around the treated area.