From Wikipedia, the free encyclopedia
The theory of recapitulation, also called the biogenetic law or embryological parallelism— often expressed in Ernst Haeckel's phrase "ontogeny recapitulates phylogeny"—is a largely discredited biological hypothesis that in developing from embryo to adult, animals go through stages resembling or representing successive stages in the evolution of their remote ancestors. While examples of embryonic stages show that molecular features of ancestral organisms exist, the theory of recapitulation itself has been viewed within the field of developmental biology as a historical side-note rather than as dogma.[1][2][3]
With different formulations, such ideas have been applied and extended to several fields and areas, including the origin of language, religion, biology, cognition and mental activities,[4] anthropology,[5] education theory[6] and developmental psychology.[7] Recapitulation theory is still considered plausible and is applied by some researchers in fields such as the study of the origin of language,[8] cognitive development,[9] behavioral development in animal species.[10]
The idea was reprised in 1720 by Giambattista Vico in his influential Scienza Nuova.[13][14][15] It was first formulated in biology in the 1790s among the German Natural philosophers,[16] after which, Marcel Danesi states, it soon gained the status of a supposed biogenetic law.[13]
The first formal formulation was proposed by Étienne Serres in 1824–26 as what became known as the "Meckel-Serres Law", it attempted to provide a link between comparative embryology and a "pattern of unification" in the organic world. It was supported by Étienne Geoffroy Saint-Hilaire and became a prominent part of his ideas which suggested that past transformations of life could have had environmental causes working on the embryo, rather than on the adult as in Lamarckism. These naturalistic ideas led to disagreements with Georges Cuvier. It was widely supported in the Edinburgh and London schools of higher anatomy around 1830, notably by Robert Edmond Grant, but was opposed by Karl Ernst von Baer's ideas of divergence, and attacked by Richard Owen in the 1830s.[17]
Ernst Haeckel attempted to synthesize the ideas of Lamarckism and Goethe's Naturphilosophie with Charles Darwin's concepts. While often seen as rejecting Darwin's theory of branching evolution for a more linear Lamarckian "biogenic law" of progressive evolution, this is not accurate: Haeckel used the Lamarckian picture to describe the ontogenetic and phylogenetic history of individual species, but agreed with Darwin about the branching of all species from one, or a few, original ancestors.[19]
Since early in the twentieth century, Haeckel's "biogenetic law" has been refuted on many fronts.[3]
Haeckel formulated his theory as "Ontogeny recapitulates phylogeny". The notion later became simply known as the recapitulation theory. Ontogeny is the growth (size change) and development (shape change) of an individual organism; phylogeny is the evolutionary history of a species. Haeckel claimed that the development of advanced species passes through stages represented by adult organisms of more primitive species.[3] Otherwise put, each successive stage in the development of an individual represents one of the adult forms that appeared in its evolutionary history.
For example, Haeckel proposed that the pharyngeal grooves between the pharyngeal arches in the neck of the human embryo not only roughly resembled gill slits of fish, but directly represented an adult "fishlike" developmental stage, signifying a fishlike ancestor. Embryonic pharyngeal slits, which form in many animals when the thin branchial plates separating pharyngeal pouches and pharyngeal grooves perforate, open the pharynx to the outside. Pharyngeal arches appear in all tetrapod embryos: in mammals, the first pharyngeal arch develops into the lower jaw (Meckel's cartilage), the malleus and the stapes. But these embryonic pharyngeal arches, grooves, pouches, and slits in human embryos can not at any stage carry out the same function as the gills of an adult fish.
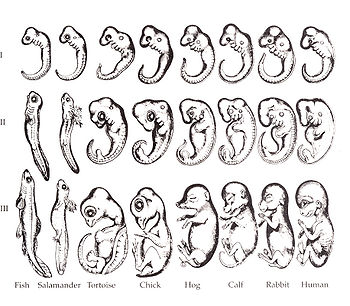
Haeckel produced several embryo drawings that often overemphasized similarities between embryos of related species. The misinformation was propagated through many biology textbooks, and popular knowledge, even today. Modern biology rejects the literal and universal form of Haeckel's theory.[20]
Haeckel's drawings were disputed by Wilhelm His, who had developed a rival theory of embryology.[21] His developed a "causal-mechanical theory" of human embryonic development.[22]
Darwin's view, that early embryonic stages are similar to the same embryonic stage of related species but not to the adult stages of these species, has been confirmed by modern evolutionary developmental biology[citation needed].
English philosopher Herbert Spencer was one of the most energetic promoters of evolutionary ideas to explain many phenomena. He compactly expressed the basis for a cultural recapitulation theory of education in the following claim, published in 1861, five years before Haeckel first published on the subject:[6] G. Stanley Hall used Haeckel's theories as the basis for his theories of child development.
The Austrian pioneer in psychoanalysis, Sigmund Freud, also favored Haeckel's doctrine. He was trained as a biologist under the influence of recapitulation theory at the time of its domination, and retained a Lamarckian outlook with justification from the recapitulation theory.[28] He also distinguished between physical and mental recapitulation, in which the differences would become an essential argument for his theory of neuroses.[28]
Taruskin also developed a variation of the motto into the pun "ontogeny recapitulates ontology" to refute the concept of "absolute music" advancing the socio-artistic theories of Carl Dalhaus. Ontology is the investigation of what exactly something is, and Taruskin asserts that an art object becomes that which society and succeeding generations made of it. For example, composer Johann Sebastian Bach's St. John Passion, composed in the 1720s, was appropriated by the Nazi regime in the 1930s for propaganda. Taruskin claims the historical development of the Passion (its ontogeny) as a work with an anti-Semitic message does, in fact, inform the work's identity (its ontology), even though that was an unlikely concern of the composer. Music or even an abstract visual artwork can not be truly autonomous ("absolute") because it is defined by its historical and social reception.[29]
With different formulations, such ideas have been applied and extended to several fields and areas, including the origin of language, religion, biology, cognition and mental activities,[4] anthropology,[5] education theory[6] and developmental psychology.[7] Recapitulation theory is still considered plausible and is applied by some researchers in fields such as the study of the origin of language,[8] cognitive development,[9] behavioral development in animal species.[10]
Origins
The earliest recorded trace of a recapitulation theory is from the Egyptian Pharaoh Psamtik I (664 – 610 BCE), who used it as a hypothesis on the origin of language.[11][12] The concept of recapitulation was first formulated outside the field of biology. It was widely held among traditional theories of the origin of language (glottology), being assumed as a premise that children's use of language gives insights on its origin and evolution.[13]The idea was reprised in 1720 by Giambattista Vico in his influential Scienza Nuova.[13][14][15] It was first formulated in biology in the 1790s among the German Natural philosophers,[16] after which, Marcel Danesi states, it soon gained the status of a supposed biogenetic law.[13]
The first formal formulation was proposed by Étienne Serres in 1824–26 as what became known as the "Meckel-Serres Law", it attempted to provide a link between comparative embryology and a "pattern of unification" in the organic world. It was supported by Étienne Geoffroy Saint-Hilaire and became a prominent part of his ideas which suggested that past transformations of life could have had environmental causes working on the embryo, rather than on the adult as in Lamarckism. These naturalistic ideas led to disagreements with Georges Cuvier. It was widely supported in the Edinburgh and London schools of higher anatomy around 1830, notably by Robert Edmond Grant, but was opposed by Karl Ernst von Baer's ideas of divergence, and attacked by Richard Owen in the 1830s.[17]
Haeckel

George Romanes's 1892 copy of Ernst Haeckel's controversial embryo drawings (this version of the figure is often attributed incorrectly to Haeckel).[18]
Ernst Haeckel attempted to synthesize the ideas of Lamarckism and Goethe's Naturphilosophie with Charles Darwin's concepts. While often seen as rejecting Darwin's theory of branching evolution for a more linear Lamarckian "biogenic law" of progressive evolution, this is not accurate: Haeckel used the Lamarckian picture to describe the ontogenetic and phylogenetic history of individual species, but agreed with Darwin about the branching of all species from one, or a few, original ancestors.[19]
Since early in the twentieth century, Haeckel's "biogenetic law" has been refuted on many fronts.[3]
Haeckel formulated his theory as "Ontogeny recapitulates phylogeny". The notion later became simply known as the recapitulation theory. Ontogeny is the growth (size change) and development (shape change) of an individual organism; phylogeny is the evolutionary history of a species. Haeckel claimed that the development of advanced species passes through stages represented by adult organisms of more primitive species.[3] Otherwise put, each successive stage in the development of an individual represents one of the adult forms that appeared in its evolutionary history.
For example, Haeckel proposed that the pharyngeal grooves between the pharyngeal arches in the neck of the human embryo not only roughly resembled gill slits of fish, but directly represented an adult "fishlike" developmental stage, signifying a fishlike ancestor. Embryonic pharyngeal slits, which form in many animals when the thin branchial plates separating pharyngeal pouches and pharyngeal grooves perforate, open the pharynx to the outside. Pharyngeal arches appear in all tetrapod embryos: in mammals, the first pharyngeal arch develops into the lower jaw (Meckel's cartilage), the malleus and the stapes. But these embryonic pharyngeal arches, grooves, pouches, and slits in human embryos can not at any stage carry out the same function as the gills of an adult fish.
Haeckel produced several embryo drawings that often overemphasized similarities between embryos of related species. The misinformation was propagated through many biology textbooks, and popular knowledge, even today. Modern biology rejects the literal and universal form of Haeckel's theory.[20]
Haeckel's drawings were disputed by Wilhelm His, who had developed a rival theory of embryology.[21] His developed a "causal-mechanical theory" of human embryonic development.[22]
Darwin's view, that early embryonic stages are similar to the same embryonic stage of related species but not to the adult stages of these species, has been confirmed by modern evolutionary developmental biology[citation needed].
Modern status
The Haeckelian form of recapitulation theory is considered defunct.[23] However, embryos do undergo a period where their morphology is strongly shaped by their phylogenetic position, rather than selective pressures.[24]"Embryos do reflect the course of evolution, but that course is far more intricate and quirky than Haeckel claimed. Different parts of the same embryo can even evolve in different directions. As a result, the Biogenetic Law was abandoned, and its fall freed scientists to appreciate the full range of embryonic changes that evolution can produce—an appreciation that has yielded spectacular results in recent years as scientists have discovered some of the specific genes that control development."[25]
Influence
Cognitive development
Although Haeckel's specific form of recapitulation theory is now discredited among biologists, the strong influence it had on social and educational theories of the late 19th century still resonates in the 21st century. Research in the late 20th century confirmed that "both biological evolution and the stages in the child’s cognitive development follow much the same progression of evolutionary stages as that suggested in the archaeological record."[9]English philosopher Herbert Spencer was one of the most energetic promoters of evolutionary ideas to explain many phenomena. He compactly expressed the basis for a cultural recapitulation theory of education in the following claim, published in 1861, five years before Haeckel first published on the subject:[6] G. Stanley Hall used Haeckel's theories as the basis for his theories of child development.
If there be an order in which the human race has mastered its various kinds of knowledge, there will arise in every child an aptitude to acquire these kinds of knowledge in the same order.... Education is a repetition of civilization in little.[26]Developmental psychologist Jean Piaget favored a weaker version of the formula, according to which ontogeny parallels phylogeny because the two are subject to similar external constraints.[27]
— Herbert Spencer
The Austrian pioneer in psychoanalysis, Sigmund Freud, also favored Haeckel's doctrine. He was trained as a biologist under the influence of recapitulation theory at the time of its domination, and retained a Lamarckian outlook with justification from the recapitulation theory.[28] He also distinguished between physical and mental recapitulation, in which the differences would become an essential argument for his theory of neuroses.[28]
Art criticism
More recently, several art historians, most prominently musicologist Richard Taruskin, have applied the term "ontogeny becomes phylogeny" to the process of creating and recasting art history, often to assert a perspective or argument. For example, the peculiar development of the works by modernist composer Arnold Schoenberg (here an "ontogeny") is generalized in many histories into a "phylogeny" – a historical development ("evolution") of Western Music toward atonal styles of which Schoenberg is a representative. Such historiographies of the "collapse of traditional tonality" are faulted by art historians as asserting a rhetorical rather than historical point about tonality's "collapse".[29]Taruskin also developed a variation of the motto into the pun "ontogeny recapitulates ontology" to refute the concept of "absolute music" advancing the socio-artistic theories of Carl Dalhaus. Ontology is the investigation of what exactly something is, and Taruskin asserts that an art object becomes that which society and succeeding generations made of it. For example, composer Johann Sebastian Bach's St. John Passion, composed in the 1720s, was appropriated by the Nazi regime in the 1930s for propaganda. Taruskin claims the historical development of the Passion (its ontogeny) as a work with an anti-Semitic message does, in fact, inform the work's identity (its ontology), even though that was an unlikely concern of the composer. Music or even an abstract visual artwork can not be truly autonomous ("absolute") because it is defined by its historical and social reception.[29]






