Commonly used protein production systems include those derived from bacteria, yeast,baculovirus/insect, mammalian cells, and more recently filamentous fungi such as Myceliophthora thermophila. When biopharmaceuticals are produced with one of these systems, process-related impurities termed host cell proteins also arrive in the final product in trace amounts.
Cell-based systems
The oldest and most widely used expression systems are cell-based and may be defined as the "combination of an expression vector,
its cloned DNA, and the host for the vector that provide a context to
allow foreign gene function in a host cell, that is, produce proteins at
a high level".Overexpression is an abnormally and excessively high level of gene expression which produces a pronounced gene-related phenotype.
There are many ways to introduce foreign DNA
to a cell for expression, and many different host cells may be used for
expression — each expression system has distinct advantages and
liabilities. Expression systems are normally referred to by the host and the DNA source or the delivery mechanism for the genetic material. For example, common hosts are bacteria (such as E.coli, B. subtilis), yeast (such as S.cerevisiae) or eukaryotic cell lines. Common DNA sources and delivery mechanisms are viruses (such as baculovirus, retrovirus, adenovirus), plasmids, artificial chromosomes and bacteriophage (such as lambda). The best expression system depends on the gene involved, for example the Saccharomyces cerevisiae is often preferred for proteins that require significant posttranslational modification. Insect or mammal
cell lines are used when human-like splicing of mRNA is required.
Nonetheless, bacterial expression has the advantage of easily producing
large amounts of protein, which is required for X-ray crystallography or nuclear magnetic resonance experiments for structure determination.
Because bacteria are prokaryotes,
they are not equipped with the full enzymatic machinery to accomplish
the required post-translational modifications or molecular folding.
Hence, multi-domain eukaryotic proteins expressed in bacteria often are
non-functional. Also, many proteins become insoluble as inclusion bodies
that are difficult to recover without harsh denaturants and subsequent
cumbersome protein-refolding.
To address these concerns, expressions systems using multiple
eukaryotic cells were developed for applications requiring the proteins
be conformed as in, or closer to eukaryotic organisms: cells of plants
(i.e. tobacco), of insects or mammalians (i.e. bovines) are transfected
with genes and cultured in suspension and even as tissues or whole
organisms, to produce fully folded proteins. Mammalian in vivo
expression systems have however low yield and other limitations
(time-consuming, toxicity to host cells,..). To combine the high
yield/productivity and scalable protein features of bacteria and yeast,
and advanced epigenetic features of plants, insects and mammalians
systems, other protein production systems are developed using
unicellular eukaryotes (i.e. non-pathogenic 'Leishmania' cells).
Bacterial systems
Escherichia coli
E. coli, one of the most popular hosts for artificial gene expression.
E. coli is one of the most widely used expression hosts, and DNA is normally introduced in a plasmid expression vector. The techniques for overexpression in E. coli
are well developed and work by increasing the number of copies of the
gene or increasing the binding strength of the promoter region so
assisting transcription.
For example, a DNA sequence for a protein of interest could be cloned or subcloned into a high copy-number plasmid containing the lac (often LacUV5) promoter, which is then transformed into the bacterium E. coli. Addition of IPTG (a lactose analog) activates the lac promoter and causes the bacteria to express the protein of interest.
E. coli strain BL21 and BL21(DE3) are two strains commonly used for protein production. As members of the B lineage, they lack lon and OmpT proteases, protecting the produced proteins from degradation. The DE3 prophage found in BL21(DE3) provides T7 RNA polymerase (driven by the LacUV5 promoter), allowing for vectors with the T7 promoter to be used instead.
Corynebacterium
Non-pathogenic species of the gram-positive Corynebacterium are used for the commercial production of various amino acids. The C. glutamicum species is widely used for producing glutamate and lysine, components of human food, animal feed and pharmaceutical products.
Expression of functionally active human epidermal growth factor has been done in C. glutamicum,
thus demonstrating a potential for industrial-scale production of human
proteins. Expressed proteins can be targeted for secretion through
either the general, secretory pathway (Sec) or the twin-arginine translocation pathway (Tat).
The non-pathogenic and gram-negative bacteria, Pseudomonas fluorescens, is used for high level production of recombinant proteins; commonly for the development bio-therapeutics and vaccines. P. fluorescens is a metabolically versatile organism, allowing for high throughput screening and rapid development of complex proteins. P. fluorescens is most well known for its ability to rapid and successfully produce high titers of active, soluble protein.
Eukaryotic systems
Yeasts
Expression systems using either S. cerevisiae or Pichia pastoris
allow stable and lasting production of proteins that are processed
similarly to mammalian cells, at high yield, in chemically defined media
of proteins.
Filamentous fungi
Filamentous fungi, especially Aspergillus and Trichoderma, but also more recently Myceliophthora thermophila C1 have been developed into expression platforms for screening and production of diverse industrial enzymes.
The expression system C1 shows a low viscosity morphology in submerged
culture, enabling the use of complex growth and production media.
Baculovirus-infected cells
Baculovirus-infected insect cells (Sf9, Sf21, High Five strains) or mammalian cells (HeLa, HEK 293) allow production of glycosylated or membrane proteins that cannot be produced using fungal or bacterial systems.
It is useful for production of proteins in high quantity. Genes are not
expressed continuously because infected host cells eventually lyse and
die during each infection cycle.
Non-lytic insect cell expression
Non-lytic
insect cell expression is an alternative to the lytic baculovirus
expression system. In non-lytic expression, vectors are transiently or
stably transfected into the chromosomal DNA of insect cells for subsequent gene expression. This is followed by selection and screening of recombinant clones.
The non-lytic system has been used to give higher protein yield and
quicker expression of recombinant genes compared to baculovirus-infected
cell expression. Cell lines used for this system include: Sf9, Sf21 from Spodoptera frugiperda cells, Hi-5 from Trichoplusia ni cells, and Schneider 2 cells and Schneider 3 cells from Drosophila melanogaster cells. With this system, cells do not lyse and several cultivation modes can be used. Additionally, protein production runs are reproducible. This system gives a homogeneous product. A drawback of this system is the requirement of an additional screening step for selecting viable clones.
Leishmania tarentolae
(cannot infect mammals) expression systems allow stable and lasting
production of proteins at high yield, in chemically defined media.
Produced proteins exhibit fully eukaryotic post-translational
modifications, including glycosylation and disulfide bond formation.
Mammalian systems
The most common mammalian expression systems are Chinese Hamsterovary (CHO) and Human embryonic kidney (HEK) cells.
Cell-free production of proteins is performed in vitro
using purified RNA polymerase, ribosomes, tRNA and ribonucleotides.
These reagents may be produced by extraction from cells or from a
cell-based expression system. Due to the low expression levels and high
cost of cell-free systems, cell-based systems are more widely used.
Proteomics is the large-scale study of proteins. Proteins are vital parts of living organisms, with many functions. The proteome
is the entire set of proteins that is produced or modified by an
organism or system. Proteomics has enabled the identification of ever
increasing numbers of protein. This varies with time and distinct
requirements, or stresses, that a cell or organism undergoes.
Proteomics is an interdisciplinary domain that has benefitted greatly
from the genetic information of various genome projects, including the Human Genome Project.
It covers the exploration of proteomes from the overall level of
protein composition, structure, and activity. It is an important
component of functional genomics.
Proteomics generally refers to the large-scale
experimental analysis of proteins and proteomes, but often is used
specifically to refer to protein purification and mass spectrometry.
History and etymology
The
first studies of proteins that could be regarded as proteomics began in
1975, after the introduction of the two-dimensional gel and mapping of
the proteins from the bacterium Escherichia coli.
The word proteome is blend of the words "protein" and "genome", and was coined by Marc Wilkins in 1994 while he was a Ph.D. student at Macquarie University. Macquarie University also founded the first dedicated proteomics laboratory in 1995.
Complexity of the problem
After genomics and transcriptomics,
proteomics is the next step in the study of biological systems. It is
more complicated than genomics because an organism's genome is more or
less constant, whereas proteomes differ from cell to cell and from time
to time. Distinct genes are expressed in different cell types, which means that even the basic set of proteins that are produced in a cell needs to be identified.
In the past this phenomenon was assessed by RNA analysis, but it was found to lack correlation with protein content. Now it is known that mRNA is not always translated into protein,
and the amount of protein produced for a given amount of mRNA depends
on the gene it is transcribed from and on the current physiological
state of the cell. Proteomics confirms the presence of the protein and
provides a direct measure of the quantity present.
Post-translational modifications
Not only does the translation from mRNA cause differences, but many
proteins also are subjected to a wide variety of chemical modifications
after translation. The most common and widely studied post translational
modifications include phosphorylation and glycosylation. Many of these
post-translational modifications are critical to the protein's function.
Phosphorylation
One such modification is phosphorylation, which happens to many enzymes and structural proteins in the process of cell signaling. The addition of a phosphate to particular amino acids—most commonly serine and threonine mediated by serine-threonine kinases, or more rarely tyrosine mediated by tyrosine kinases—causes
a protein to become a target for binding or interacting with a distinct
set of other proteins that recognize the phosphorylated domain.
Because protein phosphorylation is one of the most-studied
protein modifications, many "proteomic" efforts are geared to
determining the set of phosphorylated proteins in a particular cell or
tissue-type under particular circumstances. This alerts the scientist to
the signaling pathways that may be active in that instance.
Ubiquitination
Ubiquitin is a small protein that may be affixed to certain protein substrates by enzymes called E3 ubiquitin ligases.
Determining which proteins are poly-ubiquitinated helps understand how
protein pathways are regulated. This is, therefore, an additional
legitimate "proteomic" study. Similarly, once a researcher determines
which substrates are ubiquitinated by each ligase, determining the set
of ligases expressed in a particular cell type is helpful.
Distinct proteins are made under distinct settings
A cell may make different sets of proteins at different times or under different conditions, for example during development, cellular differentiation, cell cycle, or carcinogenesis.
Further increasing proteome complexity, as mentioned, most proteins are
able to undergo a wide range of post-translational modifications.
Therefore, a "proteomics" study may become complex very quickly,
even if the topic of study is restricted. In more ambitious settings,
such as when a biomarker
for a specific cancer subtype is sought, the proteomics scientist might
elect to study multiple blood serum samples from multiple cancer
patients to minimise confounding factors and account for experimental
noise. Thus, complicated experimental designs are sometimes necessary to account for the dynamic complexity of the proteome.
Limitations of genomics and proteomics studies
Proteomics gives a different level of understanding than genomics for many reasons:
the level of transcription of a gene gives only a rough estimate of its level of translation into a protein. An mRNA produced in abundance may be degraded rapidly or translated inefficiently, resulting in a small amount of protein.
as mentioned above, many proteins experience post-translational modifications
that profoundly affect their activities; for example, some proteins are
not active until they become phosphorylated. Methods such as phosphoproteomics and glycoproteomics are used to study post-translational modifications.
many transcripts give rise to more than one protein, through alternative splicing or alternative post-translational modifications.
many proteins form complexes with other proteins or RNA molecules, and only function in the presence of these other molecules.
protein degradation rate plays an important role in protein content.
Reproducibility. One major factor affecting reproducibility in proteomics experiments is the simultaneous elution of many more peptides than mass spectrometers can measure. This causes stochastic differences between experiments due to data-dependent acquisition
of tryptic peptides. Although early large-scale shotgun proteomics
analyses showed considerable variability between laboratories,
presumably due in part to technical and experimental differences
between laboratories, reproducibility has been improved in more recent
mass spectrometry analysis, particularly on the protein level and using
Orbitrap mass spectrometers. Notably, targeted proteomics
shows increased reproducibility and repeatability compared with shotgun
methods, although at the expense of data density and effectiveness.
Methods of studying proteins
In proteomics, there are multiple methods to study proteins. Generally, proteins may be detected by using either antibodies (immunoassays) or mass spectrometry.
If a complex biological sample is analyzed, either a very specific
antibody needs to be used in quantitative dot blot analysis (QDB), or
biochemical separation then needs to be used before the detection step,
as there are too many analytes in the sample to perform accurate
detection and quantification.
Protein detection with antibodies (immunoassays)
Antibodies to particular proteins, or to their modified forms, have been used in biochemistry and cell biology
studies. These are among the most common tools used by molecular
biologists today. There are several specific techniques and protocols
that use antibodies for protein detection. The enzyme-linked immunosorbent assay (ELISA) has been used for decades to detect and quantitatively measure proteins in samples. The western blot
may be used for detection and quantification of individual proteins,
where in an initial step, a complex protein mixture is separated using SDS-PAGE and then the protein of interest is identified using an antibody.
Modified proteins may be studied by developing an antibody specific to that modification. For example, there are antibodies that only recognize certain proteins when they are tyrosine-phosphorylated,
they are known as phospho-specific antibodies. Also, there are
antibodies specific to other modifications. These may be used to
determine the set of proteins that have undergone the modification of
interest.
Disease detection at the molecular level is driving the emerging
revolution of early diagnosis and treatment. A challenge facing the
field is that protein biomarkers for early diagnosis may be present in
very low abundance. The lower limit of detection with conventional
immunoassay technology is the upper femtomolar range (10−13 M). Digital immunoassay technology has improved detection sensitivity three logs, to the attomolar range (10−16
M). This capability has the potential to open new advances in
diagnostics and therapeutics, but such technologies have been relegated
to manual procedures that are not well suited for efficient routine use.
Antibody-free protein detection
While
protein detection with antibodies is still very common in molecular
biology, other methods have been developed as well, that do not rely on
an antibody. These methods offer various advantages, for instance they
often are able to determine the sequence of a protein or peptide, they
may have higher throughput than antibody-based, and they sometimes can
identify and quantify proteins for which no antibody exists.
Detection methods
One of the earliest methods for protein analysis has been Edman degradation (introduced in 1967) where a single peptide
is subjected to multiple steps of chemical degradation to resolve its
sequence. These early methods have mostly been supplanted by
technologies that offer higher throughput.
For the analysis of complex biological samples, a reduction of sample complexity is required. This may be performed off-line by one-dimensional or two-dimensional
separation. More recently, on-line methods have been developed where
individual peptides (in bottom-up proteomics approaches) are separated
using reversed-phase chromatography and then, directly ionized using ESI; the direct coupling of separation and analysis explains the term "on-line" analysis.
Hybrid technologies
There
are several hybrid technologies that use antibody-based purification of
individual analytes and then perform mass spectrometric analysis for
identification and quantification. Examples of these methods are the
MSIA (mass spectrometric immunoassay), developed by Randall Nelson in 1995,[20] and the SISCAPA (Stable Isotope Standard Capture with Anti-Peptide Antibodies) method, introduced by Leigh Anderson in 2004.
Current research methodologies
Fluorescence two-dimensional differential gel electrophoresis (2-D DIGE)
may be used to quantify variation in the 2-D DIGE process and establish
statistically valid thresholds for assigning quantitative changes
between samples.
Comparative proteomic analysis may reveal the role of proteins in
complex biological systems, including reproduction. For example,
treatment with the insecticide triazophos causes an increase in the
content of brown planthopper (Nilaparvata lugens (Stål)) male
accessory gland proteins (Acps) that may be transferred to females via
mating, causing an increase in fecundity (i.e. birth rate) of females.
To identify changes in the types of accessory gland proteins (Acps) and
reproductive proteins that mated female planthoppers received from male
planthoppers, researchers conducted a comparative proteomic analysis of
mated N. lugens females. The results indicated that these proteins participate in the reproductive process of N. lugens adult females and males.
Proteome analysis of Arabidopsis peroxisomes has been established as the major unbiased approach for identifying new peroxisomal proteins on a large scale.
There are many approaches to characterizing the human proteome,
which is estimated to contain between 20,000 and 25,000 non-redundant
proteins. The number of unique protein species likely will increase by
between 50,000 and 500,000 due to RNA splicing and proteolysis events,
and when post-translational modification also are considered, the total
number of unique human proteins is estimated to range in the low
millions.
In addition, the first promising attempts to decipher the proteome of animal tumors have recently been reported.
This method was used as a functional method in Macrobrachium rosenbergii protein profiling.
High-throughput proteomic technologies
Proteomics
has steadily gained momentum over the past decade with the evolution of
several approaches. Few of these are new, and others build on
traditional methods. Mass spectrometry-based methods and micro arrays
are the most common technologies for large-scale study of proteins.
Mass spectrometry and protein profiling
There are two mass spectrometry-based methods currently used for
protein profiling. The more established and widespread method uses high
resolution, two-dimensional electrophoresis to separate proteins from
different samples in parallel, followed by selection and staining of
differentially expressed proteins to be identified by mass spectrometry.
Despite the advances in 2-DE and its maturity, it has its limits as
well.
The central concern is the inability to resolve all the proteins within a
sample, given their dramatic range in expression level and differing
properties.
The second quantitative approach uses stable isotope tags to
differentially label proteins from two different complex mixtures. Here,
the proteins within a complex mixture are labeled isotopically first,
and then digested to yield labeled peptides. The labeled mixtures are
then combined, the peptides separated by multidimensional liquid
chromatography and analyzed by tandem mass spectrometry. Isotope coded
affinity tag (ICAT) reagents are the widely used isotope tags. In this
method, the cysteine residues of proteins get covalently attached to the
ICAT reagent, thereby reducing the complexity of the mixtures omitting
the non-cysteine residues.
Quantitative proteomics using stable isotopic tagging is an
increasingly useful tool in modern development. Firstly, chemical
reactions have been used to introduce tags into specific sites or
proteins for the purpose of probing specific protein functionalities.
The isolation of phosphorylated peptides has been achieved using
isotopic labeling and selective chemistries to capture the fraction of
protein among the complex mixture. Secondly, the ICAT technology was
used to differentiate between partially purified or purified
macromolecular complexes such as large RNA polymerase II pre-initiation
complex and the proteins complexed with yeast transcription factor.
Thirdly, ICAT labeling was recently combined with chromatin isolation to
identify and quantify chromatin-associated proteins. Finally ICAT
reagents are useful for proteomic profiling of cellular organelles and
specific cellular fractions.
Another quantitative approach is the accurate mass and time (AMT) tag approach developed by Richard D. Smith and coworkers at Pacific Northwest National Laboratory.
In this approach, increased throughput and sensitivity is achieved by
avoiding the need for tandem mass spectrometry, and making use of
precisely determined separation time information and highly accurate
mass determinations for peptide and protein identifications.
Protein chips
Balancing
the use of mass spectrometers in proteomics and in medicine is the use
of protein micro arrays. The aim behind protein micro arrays is to print
thousands of protein detecting features for the interrogation of
biological samples. Antibody arrays are an example in which a host of
different antibodies are arrayed to detect their respective antigens
from a sample of human blood. Another approach is the arraying of
multiple protein types for the study of properties like protein-DNA,
protein-protein and protein-ligand interactions. Ideally, the functional
proteomic arrays would contain the entire complement of the proteins of
a given organism. The first version of such arrays consisted of 5000
purified proteins from yeast deposited onto glass microscopic slides.
Despite the success of first chip, it was a greater challenge for
protein arrays to be implemented. Proteins are inherently much more
difficult to work with than DNA. They have a broad dynamic range, are
less stable than DNA and their structure is difficult to preserve on
glass slides, though they are essential for most assays. The global ICAT
technology has striking advantages over protein chip technologies.
Reverse-phased protein microarrays
This
is a promising and newer microarray application for the diagnosis,
study and treatment of complex diseases such as cancer. The technology
merges laser capture microdissection (LCM)
with micro array technology, to produce reverse phase protein
microarrays. In this type of microarrays, the whole collection of
protein themselves are immobilized with the intent of capturing various
stages of disease within an individual patient. When used with LCM,
reverse phase arrays can monitor the fluctuating state of proteome among
different cell population within a small area of human tissue. This is
useful for profiling the status of cellular signaling molecules, among a
cross section of tissue that includes both normal and cancerous cells.
This approach is useful in monitoring the status of key factors in
normal prostate epithelium and invasive prostate cancer tissues. LCM
then dissects these tissue and protein lysates were arrayed onto
nitrocellulose slides, which were probed with specific antibodies. This
method can track all kinds of molecular events and can compare diseased
and healthy tissues within the same patient enabling the development of
treatment strategies and diagnosis. The ability to acquire proteomics
snapshots of neighboring cell populations, using reverse phase
microarrays in conjunction with LCM has a number of applications beyond
the study of tumors. The approach can provide insights into normal
physiology and pathology of all the tissues and is invaluable for
characterizing developmental processes and anomalies.
Practical applications
New Drug Discovery
One
major development to come from the study of human genes and proteins
has been the identification of potential new drugs for the treatment of
disease. This relies on genome and proteome
information to identify proteins associated with a disease, which
computer software can then use as targets for new drugs. For example, if
a certain protein is implicated in a disease, its 3D structure provides
the information to design drugs to interfere with the action of the
protein. A molecule that fits the active site of an enzyme, but cannot
be released by the enzyme, inactivates the enzyme. This is the basis of
new drug-discovery tools, which aim to find new drugs to inactivate
proteins involved in disease. As genetic differences among individuals
are found, researchers expect to use these techniques to develop
personalized drugs that are more effective for the individual.
Proteomics is also used to reveal complex plant-insect
interactions that help identify candidate genes involved in the
defensive response of plants to herbivory.
Expression proteomics includes the analysis of protein expression
at larger scale. It helps identify main proteins in a particular
sample, and those proteins differentially expressed in related
samples—such as diseased vs. healthy tissue. If a protein is found only
in a diseased sample then it can be a useful drug target or diagnostic
marker. Proteins with same or similar expression profiles may also be
functionally related. There are technologies such as 2D-PAGE and mass spectrometry that are used in expression proteomics.
Biomarkers
The National Institutes of Health
has defined a biomarker as "a characteristic that is objectively
measured and evaluated as an indicator of normal biological processes,
pathogenic processes, or pharmacologic responses to a therapeutic
intervention."
Understanding the proteome, the structure and function of each
protein and the complexities of protein–protein interactions is critical
for developing the most effective diagnostic techniques and disease
treatments in the future. For example, proteomics is highly useful in
identification of candidate biomarkers (proteins in body fluids that are
of value for diagnosis), identification of the bacterial antigens that
are targeted by the immune response, and identification of possible
immunohistochemistry markers of infectious or neoplastic diseases.
An interesting use of proteomics is using specific protein
biomarkers to diagnose disease. A number of techniques allow to test for
proteins produced during a particular disease, which helps to diagnose
the disease quickly. Techniques include western blot, immunohistochemical staining, enzyme linked immunosorbent assay (ELISA) or mass spectrometry. Secretomics, a subfield of proteomics that studies secreted proteins
and secretion pathways using proteomic approaches, has recently emerged
as an important tool for the discovery of biomarkers of disease.
Proteogenomics
In proteogenomics, proteomic technologies such as mass spectrometry are used for improving gene annotations.
Parallel analysis of the genome and the proteome facilitates discovery
of post-translational modifications and proteolytic events, especially when comparing multiple species (comparative proteogenomics).
Structural proteomics
Structural
proteomics includes the analysis of protein structures at large-scale.
It compares protein structures and helps identify functions of newly
discovered genes. The structural analysis also helps to understand that
where drugs bind to proteins and also show where proteins interact with
each other. This understanding is achieved using different technologies
such as X-ray crystallography and NMR spectroscopy.
Bioinformatics for proteomics (proteome informatics)
Much
proteomics data is collected with the help of high throughput
technologies such as mass spectrometry and microarray. It would often
take weeks or months to analyze the data and perform comparisons by
hand. For this reason, biologists and chemists are collaborating with
computer scientists and mathematicians to create programs and pipeline
to computationally analyze the protein data. Using bioinformatics
techniques, researchers are capable of faster analysis and data
storage. A good place to find lists of current programs and databases is
on the ExPASy
bioinformatics resource portal. The applications of
bioinformatics-based proteomics includes medicine, disease diagnosis,
biomarker identification, and many more.
Protein identification
Mass
spectrometry and microarray produce peptide fragmentation information
but do not give identification of specific proteins present in the
original sample. Due to the lack of specific protein identification,
past researchers were forced to decipher the peptide fragments
themselves. However, there are currently programs available for protein
identification. These programs take the peptide sequences output from
mass spectrometry and microarray and return information about matching
or similar proteins. This is done through algorithms implemented by the
program which perform alignments with proteins from known databases such
as UniProt and PROSITE to predict what proteins are in the sample with a degree of certainty.
Protein structure
The biomolecular structure
forms the 3D configuration of the protein. Understanding the protein's
structure aids in identification of the protein's interactions and
function. It used to be that the 3D structure of proteins could only be
determined using X-ray crystallography and NMR spectroscopy. As of 2017, Cryo-electron microscopy
is a leading technique, solving difficulties with crystallization (in
X-ray crystallography) and conformational ambiguity (in NMR); resolution
was 2.2Å as of 2015. Now, through bioinformatics, there are computer
programs that can in some cases predict and model the structure of
proteins. These programs use the chemical properties of amino acids and
structural properties of known proteins to predict the 3D model of
sample proteins. This also allows scientists to model protein
interactions on a larger scale. In addition, biomedical engineers are
developing methods to factor in the flexibility of protein structures to
make comparisons and predictions.
Post-translational modifications
Most programs available for protein analysis are not written for proteins that have undergone post-translational modifications.
Some programs will accept post-translational modifications to aid in
protein identification but then ignore the modification during further
protein analysis. It is important to account for these modifications
since they can affect the protein's structure. In turn, computational
analysis of post-translational modifications has gained the attention of
the scientific community. The current post-translational modification
programs are only predictive.
Chemists, biologists and computer scientists are working together to
create and introduce new pipelines that allow for analysis of
post-translational modifications that have been experimentally
identified for their effect on the protein's structure and function.
Computational methods in studying protein biomarkers
One
example of the use of bioinformatics and the use of computational
methods is the study of protein biomarkers. Computational predictive
models
have shown that extensive and diverse feto-maternal protein trafficking
occurs during pregnancy and can be readily detected non-invasively in
maternal whole blood. This computational approach circumvented a major
limitation, the abundance of maternal proteins interfering with the
detection of fetal proteins,
to fetal proteomic analysis of maternal blood. Computational models can
use fetal gene transcripts previously identified in maternal whole blood to create a comprehensive proteomic network of the term neonate.
Such work shows that the fetal proteins detected in pregnant woman’s
blood originate from a diverse group of tissues and organs from the
developing fetus. The proteomic networks contain many biomarkers
that are proxies for development and illustrate the potential clinical
application of this technology as a way to monitor normal and abnormal
fetal development.
An information theoretic framework has also been introduced for biomarker discovery, integrating biofluid and tissue information.
This new approach takes advantage of functional synergy between certain
biofluids and tissues with the potential for clinically significant
findings not possible if tissues and biofluids were considered
individually. By conceptualizing tissue-biofluid as information
channels, significant biofluid proxies can be identified and then used
for guided development of clinical diagnostics. Candidate biomarkers are
then predicted based on information transfer criteria across the
tissue-biofluid channels. Significant biofluid-tissue relationships can
be used to prioritize clinical validation of biomarkers.
Emerging trends
A
number of emerging concepts have the potential to improve current
features of proteomics. Obtaining absolute quantification of proteins
and monitoring post-translational modifications are the two tasks that
impact the understanding of protein function in healthy and diseased
cells. For many cellular events, the protein concentrations do not
change; rather, their function is modulated by post-translational
modifications (PTM). Methods of monitoring PTM are an underdeveloped
area in proteomics. Selecting a particular subset of protein for
analysis substantially reduces protein complexity, making it
advantageous for diagnostic purposes where blood is the starting
material. Another important aspect of proteomics, yet not addressed, is
that proteomics methods should focus on studying proteins in the context
of the environment. The increasing use of chemical cross linkers,
introduced into living cells to fix protein-protein, protein-DNA and
other interactions, may ameliorate this problem partially. The challenge
is to identify suitable methods of preserving relevant interactions.
Another goal for studying protein is to develop more sophisticated
methods to image proteins and other molecules in living cells and real
time.
Systems biology
Advances in quantitative proteomics would clearly enable more in-depth analysis of cellular systems. Biological systems are subject to a variety of perturbations (cell cycle, cellular differentiation, carcinogenesis, environment (biophysical), etc.). Transcriptional and translational
responses to these perturbations results in functional changes to the
proteome implicated in response to the stimulus. Therefore, describing
and quantifying proteome-wide changes in protein abundance is crucial
towards understanding biological phenomenon more holistically, on the level of the entire system. In this way, proteomics can be seen as complementary to genomics, transcriptomics, epigenomics, metabolomics, and other -omics approaches in integrative analyses attempting to define biological phenotypes more comprehensively. As an example, The Cancer Proteome Atlas
provides quantitative protein expression data for ~200 proteins in over
4,000 tumor samples with matched transcriptomic and genomic data from The Cancer Genome Atlas.
Similar datasets in other cell types, tissue types, and species,
particularly using deep shotgun mass spectrometry, will be an immensely
important resource for research in fields like cancer biology, developmental and stem cell biology, medicine, and evolutionary biology.
Human plasma proteome
Characterizing
the human plasma proteome has become a major goal in the proteomics
arena, but it is also the most challenging proteomes of all human
tissues.
It contains immunoglobulin, cytokines, protein hormones, and secreted
proteins indicative of infection on top of resident, hemostatic
proteins. It also contains tissue leakage proteins due to the blood
circulation through different tissues in the body. The blood thus
contains information on the physiological state of all tissues and,
combined with its accessibility, makes the blood proteome invaluable for
medical purposes. It is thought that characterizing the proteome of
blood plasma is a daunting challenge.
The depth of the plasma proteome encompassing a dynamic range of more than 1010
between the highest abundant protein (albumin) and the lowest (some
cytokines) and is thought to be one of the main challenges for
proteomics.
Temporal and spatial dynamics further complicate the study of human
plasma proteome. The turnover of some proteins is quite faster than
others and the protein content of an artery may substantially vary from
that of a vein. All these differences make even the simplest proteomic
task of cataloging the proteome seem out of reach. To tackle this
problem, priorities need to be established. Capturing the most
meaningful subset of proteins among the entire proteome to generate a
diagnostic tool is one such priority. Secondly, since cancer is
associated with enhanced glycosylation of proteins, methods that focus
on this part of proteins will also be useful. Again: multiparameter
analysis best reveals a pathological state. As these technologies
improve, the disease profiles should be continually related to
respective gene expression changes.
Due to the above-mentioned problems plasma proteomics remained
challenging. However, technological advancements and continuous
developments seem to result in a revival of plasma proteomics as it was
shown recently by a technology called plasma proteome profiling.
Due to such technologies researchers were able to investigate
inflammation processes in mice, the heritability of plasma proteomes as
well as to show the effect of such a common life style change like
weight loss on the plasma proteome.
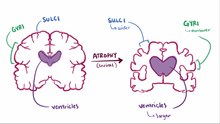
A stereotypical image of brain lateralisation - demonstrated to be false in neuroscientific research.
Neuroanatomical differences themselves exist on different scales, from neuronal densities, to the size of regions such as the planum temporale, to—at the largest scale—the torsion or "wind" in the human brain, reflected shape of the skull, which reflects a backward (posterior) protrusion of the left occipital bone and a forward (anterior) protrusion of the right frontal bone.
In addition to gross size differences, both neurochemical and
structural differences have been found between the hemispheres.
Asymmetries appear in the spacing of cortical columns, as well as
dendritic structure and complexity. Larger cell sizes are also found in
layer III of Broca's area.
The human brain has an overall leftward posterior and rightward
anterior asymmetry (or brain torque). There are particularly large
asymmetries in the frontal, temporal and occipital lobes, which increase
in asymmetry in the antero-posterior direction beginning at the central
region. Leftward asymmetry can be seen in the Heschl gyrus, parietal operculum,
Silvian fissure, left cingulate gyrus, temporo-parietal region and
planum temporale. Rightward asymmetry can be seen in the right central
sulcus (potentially suggesting increased connectivity between motor and
somatosensory cortices in the left side of the brain), lateral
ventricle, entorhinal cortex, amygdala and temporo-parieto-occipital
area. Sex-dependent brain asymmetries
are also common. For example, human male brains are more asymmetrically
lateralized than those of females. However, gene expression studies
done by Hawrylycz and colleagues and Pletikos and colleagues, were not
able to detect asymmetry between the hemispheres on the population
level.
People with autism have much more symmetrical brains than people without it.
History
In the
mid-19th century scientists first began to make discoveries regarding
lateralization of the brain, or differences in anatomy and corresponding
function between the brain's two hemispheres. Franz Gall,
a German anatomist, was the first to describe what is now known as the
Doctrine of Cerebral Localization. Gall believed that, rather than the
brain operating as a single, whole entity, different mental functions
could be attributed to different parts of the brain. He was also the
first to suggest language processing happened in the frontal lobes.
However, Gall's theories were controversial among many scientists at
the time. Others were convinced by experiments such as those conducted
by Marie-Jean-Pierre Flourens, in which he demonstrated lesions to bird brains caused irreparable damage to vital functions.
Flourens's methods, however, were not precise; the crude methodology
employed in his experiments actually caused damage to several areas of
the tiny brains of the avian models.
Paul
Broca was among the first to offer compelling evidence for localization
of function when he identified an area of the brain related to speech.
In 1861 surgeon Paul Broca
provided evidence that supported Gall's theories. Broca discovered that
two of his patients who had suffered from speech loss had similar
lesions in the same area of the left frontal lobe.
While this was compelling evidence for localization of function, the
connection to “sidedness” was not made immediately. As Broca continued
to study similar patients, he made the connection that all of the cases
involved damage to the left hemisphere, and in 1864 noted the
significance of these findings—that this must be a specialized region.
He also—incorrectly—proposed theories about the relationship of speech
areas to “handedness”.
Accordingly, some of the most famous early studies on brain asymmetry involved speech processing. Asymmetry in the Sylvian fissure
(also known as the lateral sulcus), which separates the frontal and
parietal lobes from the temporal lobe, was one of the first
incongruencies to be discovered. Its anatomical variances are related to
the size and location of two areas of the human brain that are
important for language processing, Broca's area and Wernicke's area, both in the left hemisphere.
Around the same time that Broca and Wernicke made their discoveries, neurologist Hughlings Jackson
suggested the idea of a “leading hemisphere”—or, one side of the brain
that played a more significant role in overall function—which would
eventually pave the way for understanding hemispheric “dominance” for
various processes. Several years later, in the mid-20th century,
critical understanding of hemispheric lateralization for visuospatial,
attention and perception, auditory, linguistic and emotional processing
came from patients who underwent split-brain procedures to treat disorders such as epilepsy. In split-brain patients, the corpus callosum
is cut, severing the main structure for communication between the two
hemispheres. The first modern split-brain patient was a war veteran
known as Patient W.J., whose case contributed to further understanding of asymmetry.
Brain asymmetry is not unique to humans. In addition to studies
on human patients with various diseases of the brain, much of what is
understood today about asymmetries and lateralization of function has
been learned through both invertebrate and vertebrate animal models,
including zebrafish, pigeons, rats, and many others. For example, more
recent studies revealing sexual dimorphism in brain asymmetries in the cerebral cortex and hypothalamus
of rats show that sex differences emerging from hormonal signaling can
be an important influence on brain structure and function. Work with zebrafish
has been especially informative because this species provides the best
model for directly linking asymmetric gene expression with asymmetric
morphology, and for behavioral analyses.
In humans
Lateralized functional differences and significant regions in each side of the brain and their function
The
left and right hemispheres operate the contralateral sides of the body.
Each hemisphere contains sections of all 4 lobes: the frontal lobe,
parietal lobe, temporal lobe, and occipital lobe. The two hemispheres
are separated along the mediated longitudinal fissure and are connected
by the corpus callosum which allows for communication and coordination of stimuli and information.
The corpus callosum is the largest collective pathway of white matter
tissue in the body that is made of more than 200 million nerve fibers.
The left and right hemispheres are associated with different functions
and specialize in interpreting the same data in different ways, referred
to as lateralization of the brain. The left hemisphere is associated
with language and calculations, while the right hemisphere is more
closely associated with visual-spatial recognition and facial
recognition. This lateralization of brain function
results in some specialized regions being only present in a certain
hemisphere or being dominant in one hemisphere versus the other. Some of
the significant regions included in each hemisphere are listed below.
Broca's area is located in the left hemisphere prefrontal cortex above the cingulate gyrus in the third frontal convolution.
Broca's area was discovered by Paul Broca in 1865. This area handles
speech production. Damage to this area would result in Broca aphasia
which causes the patient to become unable to formulate coherent
appropriate sentences.
Wernicke's area was discovered in 1976 by Carl Wernicke and was
found to be the site of language comprehension. Wernicke's area is also
found in the left hemisphere in the temporal lobe. Damage to this area
of the brain results in the individual losing the ability to understand
language. However, they are still able to produce sounds, words, and
sentence although they are not used in the appropriate context.
The Fusiform Face Area (FFA) is an area that has been studied to be
highly active when faces are being attended to in the visual field. A
FFA is found to be present in both hemispheres, however, studies have
found that the FFA is predominantly lateralized in the right hemisphere
where a more in-depth cognitive processing of faces is conducted. The left hemisphere FFA is associated with rapid processing of faces and their features.
Other regions and associated diseases
Some
significant regions that can present as asymmetrical in the brain can
result in either of the hemispheres due to factors such as genetics. An
example would include handedness. Handedness can result from asymmetry
in the motor cortex of one hemisphere. For right handed individuals,
since the brain operates the contralateral side of the body, they could
have a more induced motor cortex in the left hemisphere.
Several diseases have been found to exacerbate brain asymmetries
that are already present in the brain. Researchers are starting to look
into the effect and relationship of brain asymmetries to diseases such
as schizophrenia and dyslexia.
Schizophrenia is a complex long-term mental disorder that causes
hallucinations, delusions and a lack of concentration, thinking, and
motivation in an individual. Studies have found that individuals with
schizophrenia have a lack in brain asymmetry thus reducing the
functional efficiency of affected regions such as the frontal lobe.
Conditions include leftward functional hemispheric lateralization,
loss of laterality for language comprehension, a reduction in
gyrification, brain torsion etc.
As studied earlier, language is usually dominant in the left
hemisphere. Developmental language disorders, such as dyslexia, have
been researched using brain imaging techniques to understand the
neuronal or structural changes associated with the disorder. Past
research has exhibited that hemispheric asymmetries that are usually
found in healthy adults such as the size of the temporal lobe is not
present in adult patients with dyslexia. In conjunction, past research
has exhibited that patients with dyslexia lack a lateralization of
language in their brain compared to healthy patients. Instead patients
with dyslexia showed to have a bilateral hemispheric dominance for
language.
Lateralization
of function and asymmetry in the human brain continues to propel a
popular branch of neuroscientific and psychological inquiry.
Technological advancements for brain mapping have enabled researchers to
see more parts of the brain more clearly, which has illuminated
previously undetected lateralization differences that occur during
different life stages.
As more information emerges, researchers are finding insights into how
and why early human brains may have evolved the way that they did to
adapt to social, environmental and pathological changes. This
information provides clues regarding plasticity, or how different parts
of the brain can sometimes be recruited for different functions.
Continued study of brain asymmetry also contributes to the
understanding and treatment of complex diseases. Neuroimaging in
patients with Alzheimer's disease,
for example, shows significant deterioration in the left hemisphere,
along with a rightward hemispheric dominance—which could relate to
recruitment of resources to that side of the brain in the face of damage
to the left. These hemispheric changes have been connected to performance on memory tasks.
As has been the case in the past, studies on language processing
and the implications of left- and right- handedness also dominate
current research on brain asymmetry.
Evolutionary taxonomy, evolutionary systematics or Darwinian classification is a branch of biological classification that seeks to classify organisms using a combination of phylogenetic relationship (shared descent), progenitor-descendant relationship (serial descent), and degree of evolutionary change. This type of taxonomy may consider whole taxa rather than single species, so that groups of species can be inferred as giving rise to new groups. The concept found its most well-known form in the modern evolutionary synthesis of the early 1940s.
Evolutionary taxonomy differs from strict pre-Darwinian Linnaean taxonomy (producing orderly lists only), in that it builds evolutionary trees. While in phylogenetic nomenclature
each taxon must consist of a single ancestral node and all its
descendants, evolutionary taxonomy allows for groups to be excluded from
their parent taxa (e.g. dinosaurs are not considered to include birds, but to have given rise to them), thus permitting paraphyletic taxa.
Origin of evolutionary taxonomy
Jean-Baptiste Lamarck's 1815 diagram showing branching in the course of invertebrate evolution
Following the appearance of On the Origin of Species, Tree of Life representations became popular in scientific works. In On the Origin of Species,
the ancestor remained largely a hypothetical species; Darwin was
primarily occupied with showing the principle, carefully refraining from
speculating on relationships between living or fossil organisms and
using theoretical examples only. In contrast, Chambers had proposed specific hypotheses, the evolution of placental mammals from marsupials, for example.
Following Darwin's publication, Thomas Henry Huxley used the fossils of Archaeopteryx and Hesperornis to argue that the birds are descendants of the dinosaurs. Thus, a group of extant
animals could be tied to a fossil group. The resulting description,
that of dinosaurs "giving rise to" or being "the ancestors of" birds,
exhibits the essential hallmark of evolutionary taxonomic thinking.
The past three decades have seen a dramatic increase in the use
of DNA sequences for reconstructing phylogeny and a parallel shift in
emphasis from evolutionary taxonomy towards Hennig's 'phylogenetic
systematics'.
Efforts in combining modern methods of cladistics, phylogenetics, and
DNA analysis with classical views of taxonomy have recently appeared.
Certain authors have found that phylogenetic analysis is acceptable
scientifically as long as paraphyly at least for certain groups is
allowable. Such a stance is promoted in papers by Tod F. Stuessy
and others. A particularly strict form of evolutionary systematics has
been presented by Richard H. Zander in a number of papers, but
summarized in his "Framework for Post-Phylogenetic Systematics".
Briefly, Zander's pluralistic systematics is based on the
incompleteness of each of the theories: A method that cannot falsify a
hypothesis is as unscientific as a hypothesis that cannot be falsified.
Cladistics generates only trees of shared ancestry, not serial ancestry.
Taxa evolving seriatim cannot be dealt with by analyzing shared
ancestry with cladistic methods. Hypotheses such as adaptive radiation
from a single ancestral taxon cannot be falsified with cladistics.
Cladistics offers a way to cluster by trait transformations but no
evolutionary tree can be entirely dichotomous. Phylogenetics posits
shared ancestral taxa as causal agents for dichotomies yet there is no
evidence for the existence of such taxa. Molecular systematics uses DNA
sequence data for tracking evolutionary changes, thus paraphyly and
sometimes phylogenetic polyphyly signal ancestor-descendant
transformations at the taxon level, but otherwise molecular
phylogenetics makes no provision for extinct paraphyly. Additional
transformational analysis is needed to infer serial descent.
Cladogram of the moss genus Didymodon showing taxon transformations. Colors denote dissilient groups.
The
Besseyan cactus or commagram is the best evolutionary tree for showing
both shared and serial ancestry. First, a cladogram or natural key is
generated. Generalized ancestral taxa are identified and specialized
descendant taxa are noted as coming off the lineage with a line of one
color representing the progenitor through time. A Besseyan cactus or
commagram is then devised that represents both shared and serial
ancestry. Progenitor taxa may have one or more descendant taxa. Support
measures in terms of Bayes factors may be given, following Zander's
method of transformational analysis using decibans.
Cladistic analysis groups taxa by shared traits but incorporates a
dichotomous branching model borrowed from phenetics. It is essentially a
simplified dichotomous natural key, although reversals are tolerated.
The problem, of course, is that evolution is not necessarily
dichotomous. An ancestral taxon generating two or more descendants
requires a longer, less parsimonious tree. A cladogram node summarizes
all traits distal to it, not of any one taxon, and continuity in a
cladogram is from node to node, not taxon to taxon. This is not a model
of evolution, but is a variant of hierarchical cluster analysis (trait
changes and non-ultrametric branches. This is why a tree based solely on
shared traits is not called an evolutionary tree but merely a cladistic
tree. This tree reflects to a large extent evolutionary relationships
through trait transformations but ignores relationships made by
species-level transformation of extant taxa.
A Besseyan cactus evolutionary tree of the moss genus Didymodon
with generalized taxa in color and specialized descendants in white.
Support measures are given in terms of Bayes factors, using deciban
analysis of taxon transformation. Only two progenitors are considered
unknown shared ancestors.
Phylogenetics attempts to
inject a serial element by postulating ad hoc, undemonstrable shared
ancestors at each node of a cladistic tree. There are in number, for a
fully dichotomous cladogram, one less invisible shared ancestor than the
number of terminal taxa. We get, then, in effect a dichotomous natural
key with an invisible shared ancestor generating each couplet. This
cannot imply a process-based explanation without justification of the
dichotomy, and supposition of the shared ancestors as causes. The
cladistic form of analysis of evolutionary relationships cannot falsify
any genuine evolutionary scenario incorporating serial transformation,
according to Zander.
Zander has detailed methods for generating support measures for molecular serial descent
and for morphological serial descent using Bayes factors and sequential
Bayes analysis through Turing deciban or Shannon informational bit
addition.
The Tree of Life
Evolution of the vertebrates at class level, width of spindles indicating number of families. Spindle diagrams are often used in evolutionary taxonomy.
As more and more fossil groups were found and recognized in the late 19th and early 20th century, palaeontologists worked to understand the history of animals through the ages by linking together known groups. The Tree of life was slowly being mapped out, with fossil groups taking up their position in the tree as understanding increased.
These groups still retained their formal Linnaean taxonomic ranks. Some of them are paraphyletic
in that, although every organism in the group is linked to a common
ancestor by an unbroken chain of intermediate ancestors within the
group, some other descendants of that ancestor lie outside the group.
The evolution and distribution of the various taxa through time is
commonly shown as a spindle diagram (often called a Romerogram after the American palaeontologist Alfred Romer)
where various spindles branch off from each other, with each spindle
representing a taxon. The width of the spindles are meant to imply the
abundance (often number of families) plotted against time.
Vertebrate palaeontology
had mapped out the evolutionary sequence of vertebrates as currently
understood fairly well by the closing of the 19th century, followed by a
reasonable understanding of the evolutionary sequence of the plant kingdom
by the early 20th century. The tying together of the various trees into
a grand Tree of Life only really became possible with advancements in microbiology and biochemistry in the period between the World Wars.
Terminological difference
The two approaches, evolutionary taxonomy and the phylogenetic systematics derived from Willi Hennig, differ in the use of the word "monophyletic". For evolutionary systematicists, "monophyletic" means only that a group is derived from a single common ancestor. In phylogenetic nomenclature, there is an added caveat that the ancestral species and all descendants should be included in the group. The term "holophyletic" has been proposed for the latter meaning. As an example, amphibians
are monophyletic under evolutionary taxonomy, since they have arisen
from fishes only once. Under phylogenetic taxonomy, amphibians do not
constitute a monophyletic group in that the amniotes (reptiles, birds and mammals) have evolved from an amphibian ancestor and yet are not considered amphibians. Such paraphyletic groups are rejected in phylogenetic nomenclature, but are considered a signal of serial descent by evolutionary taxonomists.