| Ichneumon wasps | |
|---|---|

| |
| Diphyus sp., Rhône (France) | |

| |
| Anomaloninae, (Tanzania) | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Suborder: | Apocrita |
| Superfamily: | Ichneumonoidea |
| Family: | Ichneumonidae Latreille, 1802 |
| Subfamilies | |
|
See below
| |
The Ichneumonidae, also known as the ichneumon wasps or ichneumonids, is a parasitoid wasp family within the insect order Hymenoptera. This insect family, which so famously stirred skeptic thoughts in Charles Darwin, is among the most species-rich branches of the tree of life. At the same time, it is one of the groups for which our knowledge most severely lags behind their actual diversity. The roughly 25,000 species described today probably represent less than a quarter of their true richness, but reliable estimates are lacking, as is much of the most basic knowledge about their ecology, distribution and evolution. Ichneumonid wasps, with very few exceptions, attack the immature stages of holometabolous insects and spiders, eventually killing their hosts. They thus fulfill an important role as regulators of insect populations, both in natural and semi-natural systems, making them promising agents for biological control.
The most commonly recognized wasps are boldly colored social wasps whose females have venomous stings. They are in a separate clade: Aculeata. In contrast, ichneumonids have ovipositors instead of stingers, and they are all solitary. They use their ovipositors to lay eggs on or in the body of their prey, and the eggs hatch into carnivorous larvae that eat and kill the host.
The distribution of the ichneumonids was traditionally considered an exception to the common latitudinal gradient in species diversity, since the family was thought to be at its most species-rich in the temperate zone instead of the tropics, but numerous new tropical species have now been discovered.
Etymology and history
Insects in the family Ichneumonidae are commonly called ichneumon wasps, or ichneumonids. However, the term ichneumon wasps can refer specically to the genus Ichneumon within the Ichneumonidae and thus cause confusion. A group of ichneumonid specialists have proposed Darwin wasps as a better vernacular name for the family. Less exact terms are ichneumon flies (they are not closely related to true flies) and scorpion wasps due to the extreme lengthening and curving of the abdomen (scorpions are arachnids, not insects).
The name is derived from Latin 'ichneumon', from Ancient Greek
ἰχνεύμων (ikhneúmōn, "tracker"), from ἴχνος (íkhnos, "track, footstep").
The name first appeared in Aristotle's "History of Animals",
c. 343 BC. Aristotle noted that the ichneumon preys upon spiders, is a
wasp smaller than ordinary wasps, and carries its prey to a hole which
they lay their larvae inside, and that they seal the hole with mud. Aristotle's writing, however, more accurately describes the mud daubers than the true ichneumon wasps, which do not construct mud nests and do not sting.
Description
Adult ichneumonids superficially resemble other wasps. They have a slender waist, two pairs of wings, a pair of large compound eyes on the side of the head and three ocelli on top of the head. Their size varies considerably from a few millimetres to seven or more centimetres.
The ichneumonids have more antennal segments than typical, aculeate wasps (Aculeata: Vespoidea and Apoidea):
ichneumonids typically possess 16 or more, while most other wasps have
13 or fewer. Unlike the aculeate wasps, which sting in defense and do
not pass their eggs along the stinger, ichneumonid females have an ovipositor (homologous
to the stinger) which they use to lay eggs inside or on their host.
Ichneumonids generally inject venom along with the egg, but only larger
species (some in the genera Netelia and Ophion)
with relatively short ovipositors use the ovipositor in defense. Males
do not possess stingers or ovipositors in either lineage.
 Head (Ichneumon xanthorius). Antennae with many segments
Head (Ichneumon xanthorius). Antennae with many segments


.jpg)

_female.jpg)
Ichneumonids are distinguished from their sister group Braconidae mainly on the basis of wing venation. The fore wing of 95% of ichneumonids has vein 2m-cu (in the Comstock–Needham system),
which is absent in braconids. Vein 1rs-m of the fore wing is absent in
all ichneumonids, but is present in 85% of braconids. In the hind wing
of ichneumonids, vein rs-m joins Rs apical to (or rarely opposite) the
split between veins Rs and R1. In braconids, vein rs-m joins basal to
this split. The taxa also differ in the structure of the metasoma: about
90% of ichneumonids have a flexible suture between tergites 2 and 3,
whereas these tergites are fused in braconids (though the suture is
secondarily flexible in Aphidiinae).
 |
Ichneumonid fore wing (Syzecteus sp.). The presence of vein 2m-cu and absence of vein 1Rs+M distinguish the wing from that of braconids.
|
 |
Ichneumonid hind wing. Vein rs-m joins Rs after the split between veins Rs and R1.
|
 |
Braconid hind wing. Vein rs-m joins Rs before the split between veins Rs and R1.
|
Distribution
Ichneumonids
are found on all continents with the exception of Antarctica. They
inhabit virtually all terrestrial habitats, wherever there are suitable
invertebrate hosts.
The distribution of ichneumonid species richness is subject to
ongoing debate. Long believed to be rare in the tropics, and at its most
species rich in the temperate region, the family became a classic
textbook example of an 'exceptional' latitudinal diversity gradient. Recently this belief has been questioned, after the discovery of numerous new tropical species.
Reproduction and diet
Some ichneumonid species lay their eggs in the ground, but most inject them either directly into their host's
body or on its surface. After hatching, the ichneumonid larva consumes
its still living host. The most common hosts are larvae or pupae of Lepidoptera, Coleoptera and Hymenoptera. For example, a species of ichneumonid has been found to lay eggs in African sugarcane borer larva, a moth common in sub-Saharan Africa. Ichneumonids are also considered a primary enemy of the arctic woolly bear moth. Some species in the subfamily Pimplinae also parasitise spiders. Hyperparasitoids such as Mesochorinae
oviposit inside the larvae of other ichneumonoids. The hosts of many
species are unknown; host information has been summed up by e.g. Aubert, Perkins. and Townes.
Ichneumonids use both idiobiont and koinobiont strategies.
Idiobionts paralyze their host and prevent it from moving or growing.
Koinobionts allow their host to continue to grow and develop. In both
strategies, the host typically dies after some weeks, after which the
ichneumonid larva emerges and pupates.
Adult ichneumonids feed on a diversity of foods, including plant
sap, nectar and other insects. They spend much of their active time
searching, either for hosts (female ichneumonids) or for emerging
females (male ichneumonids).
The predation pressure exerted by ichneumonids can be tremendous, and
they are often one of the major regulators of invertebrate populations.
It is quite common for 10-20% or more of a host's population to be
parasitised (though reported parasitism rates often include
non-ichneumonid parasitoids).
 Zatypota albicoxa laying egg on a spider
Zatypota albicoxa laying egg on a spider
- Itoplectis maculator laying eggs in moth cocoons
- Rhyssa persuasoria laying eggs in dead wood, parasitising larvae of beetles or sawflies
- Therion circumflexum drinking from damaged edge of leaf
 Mating ichneumonids
Mating ichneumonids
 Larva of Acrodactyla quadrisculpta parasitising spider
Larva of Acrodactyla quadrisculpta parasitising spider
 Campoplegine pupa, with empty skin of caterpillar it parasitised above it
Campoplegine pupa, with empty skin of caterpillar it parasitised above it
 Hercus fontinalis larvae feeding on caterpillar
Hercus fontinalis larvae feeding on caterpillar





_parasitized_by_Acrodactyla_quadrisculpta_larva_(RMNH.INS.593867)_-_BDJ.1.e992.jpg)

Taxonomy and systematics

An ichneumonid caught in amber 15-20 million years ago.
The taxonomy of the ichneumonids is still poorly known. The family is
highly diverse, containing 24,000 described species. Approximately
60,000 species are estimated to exist worldwide, though some estimates
place this number at over 100,000. They are severely undersampled, and
studies of their diversity typically produce very high numbers of
species which are represented by only a single individual.
Due to the high diversity, the existence of numerous small and hard to
identify species, and the majority of species being undiscovered, it has
proven difficult to resolve the phylogeny of the ichneumonids. Even the relationships between subfamilies are unclear. The sheer diversity also means DNA sequence data is only available for a tiny fraction of the species, and detailed cladistic studies require major computing capacity.
Extensive catalogues of the ichneumonids include those by Aubert, Gauld, Perkins, and Townes.
Due to the taxonomic difficulties involved, however, their
classifications and terminology are often confusingly contradictory.
Several prominent authors have gone as far as to publish major reviews
that defy the International Code of Zoological Nomenclature.
The large number of species in Ichneumonidae may be due to the
evolution of parasitoidism in hymenoptera, which occurred approximately
247 million years ago. Ichneumonidae is the basal branch of Apocrita,
the lineage in which parasitoidism in hymenoptera evolved, and some
ichneumonids are thought to have been in stasis for millions of years
and closely resemble the common ancestor in which parasitoidism evolved.
This common ancestor was likely an Ectoparasitoid woodwasp that parasitized wood-boring beetle larvae in trees. The family has existed since at least the Early Cretaceous (ca. 125 mya), but may have appeared some time before. It diversified during the Oligocene.
Subfamilies
In 1999, the ichneumonids were divided into 39 subfamilies, whose names and definitions have varied considerably. Masoninae were added in 2019. The phylogenetic relationships between the subfamilies are still unclear.
- Acaenitinae
- Adelognathinae
- Agriotypinae
- Anomaloninae (= Anomalinae)
- Banchinae
- Brachycyrtinae (sometimes included in Labeninae)
- Campopleginae (= Porizontinae)
- Collyriinae
- Cremastinae
- Cryptinae (= Gelinae, Hemitelinae, Phygadeuontinae)
- Ctenopelmatinae (= Scolobatinae)
- Cylloceriinae (sometimes included in Microleptinae)
- Diacritinae (sometimes included in Pimplinae)
- Diplazontinae
- Eucerotinae (sometimes included in Tryphoninae)
- Hybrizontinae (= Paxylommatinae) (sometimes placed in own family)
- Ichneumoninae
- Labeninae (= Labiinae)
- Labenopimplinae (extinct)
- Lycorininae (sometimes included in Banchinae)
- Masoninae
- Mesochorinae
- Metopiinae
- Microleptinae
- Neorhacodinae (sometimes included in Banchinae)
- Nesomesochorinae
- Ophioninae
- Orthocentrinae (sometimes included in Microleptinae)
- Orthopelmatinae
- Oxytorinae
- Pedunculinae
- Phrudinae
- Pimplinae (= Ephialtinae)
- Poemeniinae (sometimes included in Pimplinae)
- Rhyssinae (sometimes included in Pimplinae)
- Sisyrostolinae (sometimes included in Phrudinae)
- Stilbopinae (sometimes included in Banchinae)
- Tatogastrinae (sometimes included in Microleptinae)
- Tersilochinae
- Tryphoninae
- Xanthocryptus novozealandicus
- Xoridinae
Famous ichneumonologists
Famous ichneumonologists include:
- Jacques Aubert
- Carl Gustav Alexander Brischke
- Peter Cameron
- Arnold Förster
- Johann Ludwig Christian Gravenhorst
- Alexander Henry Haliday
- Gerd Heinrich
- August Emil Holmgren
- Joseph Kriechbaumer
- Thomas Ansell Marshall
- Henry Keith Townes
- David Wahl
- Constantin Wesmael
Darwin and the Ichneumonidae
The
apparent cruelty of the ichneumonids troubled philosophers,
naturalists, and theologians in the 19th century, who found the
parasitoid life style inconsistent with the notion of a world created by
a loving and benevolent God. Charles Darwin
found the example of the Ichneumonidae so troubling that it contributed
to his increasing doubts about the nature and existence of a Creator.
In an 1860 letter to the American naturalist Asa Gray, Darwin wrote:
I own that I cannot see as plainly as others do, and as I should wish to do, evidence of design and beneficence on all sides of us. There seems to me too much misery in the world. I cannot persuade myself that a beneficent and omnipotent God would have designedly created the Ichneumonidae with the express intention of their feeding within the living bodies of Caterpillars, or that a cat should play with mice.
Some more recent researchers have disagreed with this, citing the
important role fulfilled by Ichneumonidae as regulators of insect
populations, both in natural and semi-natural systems. This makes them
promising agents for biological control.
Morphology
 Megarhyssa greenei female
Megarhyssa greenei female
 Morphology of the head and its processes: (А) head capsule; (В) antenna; (С) mandible
Morphology of the head and its processes: (А) head capsule; (В) antenna; (С) mandible
 Morphology of the thorax (D)
Morphology of the thorax (D)
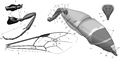 Morphology of the abdomen and processes of the thorax: (E) front wing; (F) leg III; (G) abdomen of female
Morphology of the abdomen and processes of the thorax: (E) front wing; (F) leg III; (G) abdomen of female










_(cropped).jpg)
.jpg)

.jpg)
.jpg)
.jpg)



