From Wikipedia, the free encyclopedia
The 19th century ideas of
natural selection and
Mendelian genetics were put together with
population genetics, early in the twentieth century. The modern synthesis also addressed the relationship between the broad-scale changes of
macroevolution seen by
palaeontologists and the small-scale
microevolution of local
populations of living organisms. The synthesis was defined differently by its founders, with
Ernst Mayr in 1959,
G. Ledyard Stebbins in 1966 and
Theodosius Dobzhansky
in 1974 offering differing numbers of basic postulates, though they all
included natural selection, working on heritable variation supplied by
mutation. Other major figures in the synthesis included
E. B. Ford,
Bernhard Rensch,
Ivan Schmalhausen, and
George Gaylord Simpson. An early event in the modern synthesis was
R. A. Fisher's 1918 paper on mathematical population genetics, but
William Bateson, and separately
Udny Yule, were already starting to show how Mendelian genetics could work in evolution in 1902.
Developments leading up to the synthesis
Darwin's pangenesis theory. Every part of the body emits tiny gemmules which migrate to the gonads
and contribute to the next generation via the fertilised egg. Changes
to the body during an organism's life would be inherited, as in Lamarckism.
Darwin's evolution by natural selection, 1859
The eclipse of Darwinism, 1880s onwards
From the 1880s onward, there was a widespread belief among biologists that Darwinian evolution was in deep trouble. This
eclipse of Darwinism (in
Julian Huxley's
phrase) grew out of the weaknesses in Darwin's account, written with an
incorrect view of inheritance. Darwin himself believed in
blending inheritance, which implied that any new variation, even if beneficial, would be weakened by 50% at each generation, as the engineer
Fleeming Jenkin correctly noted in 1868.
This in turn meant that small variations would not survive long enough
to be selected. Blending would therefore directly oppose natural
selection. In addition, Darwin and others considered Lamarckian
inheritance of acquired characteristics entirely possible, and Darwin's
1868 theory of
pangenesis,
with contributions to the next generation (gemmules) flowing from all
parts of the body, actually implied Lamarckism as well as blending.
Weismann's germ plasm, 1892
August Weismann's idea, set out in his 1892 book
Das Keimplasma: eine Theorie der Vererbung (The Germ Plasm: a Theory of Inheritance), was that the hereditary material, which he called the
germ plasm, and the rest of the body (the
soma)
had a one-way relationship: the germ-plasm formed the body, but the
body did not influence the germ-plasm, except indirectly in its
participation in a population subject to natural selection. If correct,
this made Darwin's pangenesis wrong, and Lamarckian inheritance
impossible. His experiment on mice, cutting off their tails and showing
that their offspring had normal tails, demonstrated that inheritance was
'hard'. He argued strongly and dogmatically
for Darwinism and against Lamarckism, polarizing opinions among other
scientists. This increased anti-Darwinian feeling, contributing to its
eclipse.
Disputed beginnings
Genetics, mutationism and biometrics, 1900–1918
While carrying out breeding experiments to clarify the mechanism of inheritance in 1900,
Hugo de Vries and
Carl Correns independently rediscovered
Gregor Mendel's work. News of this reached
William Bateson in
England, who reported on the paper during a presentation to the
Royal Horticultural Society in May 1900. In
Mendelian inheritance,
the contributions of each parent retain their integrity rather than
blending with the contribution of the other parent. In the case of a
cross between two true-breeding varieties such as Mendel's round and
wrinkled peas, the first-generation offspring are all alike, in this
case all round. Allowing these to cross, the original characteristics
reappear (segregation): about 3/4 of their offspring are round, 1/4
wrinkled. There is a discontinuity between the appearance of the
offspring; de Vries coined the term
allele for a variant form of an
inherited characteristic.
This reinforced a major division of thought, already present in the
1890s, between gradualists who followed Darwin, and saltationists such
as Bateson.
The two schools were the Mendelians, such as Bateson and de
Vries, who favored mutationism, evolution driven by mutation, based on
genes whose alleles
segregated discretely like Mendel's peas; and the
biometric school, led by
Karl Pearson and
Walter Weldon.
The biometricians argued vigorously against mutationism, saying that
empirical evidence indicated that variation was continuous in most
organisms, not discrete as Mendelism seemed to predict; they wrongly
believed that Mendelism inevitably implied evolution in discontinuous
jumps.
A traditional view is that the biometricians and the Mendelians
rejected natural selection and argued for their separate theories for 20
years, the debate only resolved by the development of population
genetics.
A more recent view is that Bateson, de Vries,
Thomas Hunt Morgan and
Reginald Punnett
had by 1918 formed a synthesis of Mendelism and mutationism. The
understanding achieved by these geneticists spanned the action of
natural selection on alleles (alternative forms of a gene), the
Hardy-Weinberg equilibrium,
the evolution of continuously-varying traits (like height), and the
probability that a new mutation will become fixed. In this view, the
early geneticists accepted natural selection but rejected Darwin's
non-Mendelian ideas about variation and heredity, and the synthesis
began soon after 1900.
The traditional claim that Mendelians rejected the idea of continuous
variation is false; as early as 1902, Bateson and Saunders wrote that
"If there were even so few as, say, four or five pairs of possible
allelomorphs, the various homo- and hetero-zygous combinations might, on
seriation, give so near an approach to a continuous curve, that the
purity of the elements would be unsuspected". Also in 1902, the statistician
Udny Yule
showed mathematically that given multiple factors, Mendel's theory
enabled continuous variation. Yule criticized Bateson's approach as
confrontational, but failed to prevent the Mendelians and the biometricians from falling out.
Castle's hooded rats, 1911
Starting in 1906,
William Castle carried out a long study of the effect of selection on coat colour in
rats. The
piebald or hooded pattern was
recessive
to the grey wild type. He crossed hooded rats with the black-backed
Irish type, and then back-crossed the offspring with pure hooded rats.
The dark stripe on the back was bigger. He then tried selecting
different groups for bigger or smaller stripes for 5 generations, and
found that it was possible to change the characteristics way beyond the
initial range of variation. This effectively refuted de Vries's claim
that continuous variation was caused by the environment and could not be
inherited. By 1911 Castle noted that the results could be explained by
Darwinian selection on heritable variation of a sufficient number of
Mendelian genes.
Morgan's fruit flies, 1912
Thomas Hunt Morgan began his career in genetics as a saltationist,
and started out trying to demonstrate that mutations could produce new
species in fruit flies. However, the experimental work at his lab with
the fruit fly,
Drosophila melanogaster
demonstrated that rather than creating new species in a single step,
mutations increased the supply of genetic variation in the population.
By 1912, after years of work on the genetics of fruit flies, Morgan
showed that these insects had many small Mendelian factors (discovered
as mutant flies) on which Darwinian evolution could work as if variation
was fully continuous. The way was open for geneticists to conclude that
Mendelism supported Darwinism.
An obstruction: Woodger's positivism, 1929
The
theoretical biologist and
philosopher of biology Joseph Henry Woodger led the introduction of
positivism into biology with his 1929 book
Biological Principles. He saw a mature
science as being characterized by a framework of
hypotheses that could be verified by facts established by
experiments. He criticized the traditional
natural history style of
biology, including the study of
evolution, as immature science, since it relied on
narrative. Woodger set out to play for biology the role of
Robert Boyle's 1661
Sceptical Chymist, intending to convert the subject into a formal, unified science, and ultimately, following the
Vienna Circle of logical positivists like
Otto Neurath and
Rudolf Carnap,
to reduce biology to physics and chemistry. His efforts stimulated the biologist
J. B. S. Haldane
to push for the axiomatization of biology, and by influencing thinkers
such as Huxley, helped to bring about the modern synthesis.
The positivist climate made natural history unfashionable, and in
America, research and university-level teaching on evolution declined
almost to nothing by the late 1930s. The Harvard physiologist
William John Crozier told his students that evolution was not even a science: "You can't experiment with two million years!"
Events in the synthesis
Fisher and Haldane's mathematical population genetics, 1918–1930
During the 1920s,
a series of papers by
J. B. S. Haldane applied mathematical analysis to real-world examples of natural selection, such as the
evolution of industrial melanism in peppered moths. Haldane established that natural selection could work even faster than Fisher had assumed.
Both workers, and others such as Dobzhansky and Wright, explicitly
intended to bring biology up to the philosophical standard of the
physical sciences, making it firmly based in mathematical modelling, its
predictions confirmed by experiment. Natural selection, once considered
hopelessly unverifiable speculation about history, was becoming
predictable, measurable, and testable.
De Beer's embryology, 1930
The traditional view is that
developmental biology played little part in the modern synthesis, but in his 1930 book
Embryos and Ancestors, the evolutionary embryologist
Gavin de Beer anticipated
evolutionary developmental biology by showing that evolution could occur by
heterochrony, such as in
the retention of juvenile features in the adult. This, de Beer argued, could cause apparently sudden changes in the
fossil record,
since embryos fossilize poorly. As the gaps in the fossil record had
been used as an argument against Darwin's gradualist evolution, de
Beer's explanation supported the Darwinian position.
However, despite de Beer, the modern synthesis largely ignored embryonic
development to explain the form of organisms, since population genetics
appeared to be an adequate explanation of how forms evolved.
Wright's adaptive landscape, 1932
The population geneticist
Sewall Wright focused on combinations of genes that interacted as complexes, and the effects of
inbreeding on small relatively isolated populations, which could be subject to
genetic drift. In a 1932 paper, he introduced the concept of an
adaptive landscape
in which phenomena such as cross breeding and genetic drift in small
populations could push them away from adaptive peaks, which would in
turn allow natural selection to push them towards new adaptive peaks.
Wright's model would appeal to field naturalists such as Theodosius
Dobzhansky and Ernst Mayr who were becoming aware of the importance of
geographical isolation in real world populations. The work of Fisher, Haldane and Wright helped to found the discipline of theoretical population genetics.
Dobzhansky's evolutionary genetics, 1937
Theodosius Dobzhansky, an emigrant from the
Soviet Union to the
United States,
who had been a postdoctoral worker in Morgan's fruit fly lab, was one
of the first to apply genetics to natural populations. He worked mostly
with
Drosophila pseudoobscura.
He says pointedly: "Russia has a variety of climates from the Arctic to
sub-tropical... Exclusively laboratory workers who neither possess nor
wish to have any knowledge of living beings in nature were and are in a
minority." Not surprisingly, there were other
Russian geneticists with similar ideas, though for some time their work was known to only a few in the
West. His 1937 work
Genetics and the Origin of Species
was a key step in bridging the gap between population geneticists and
field naturalists. It presented the conclusions reached by Fisher,
Haldane, and especially Wright in their highly mathematical papers in a
form that was easily accessible to others.
Further, Dobzhansky asserted that evolution was based on material
genes, arranged in a string on physical hereditary structures, the
chromosomes, and
linked
more or less strongly to each other according to their physical
distances from each other on the chromosomes. As with Haldane and
Fisher, Dobzhansky's "evolutionary genetics" was a genuine science, now unifying cell biology, genetics, and both micro- and macroevolution.
His work emphasized that real world populations had far more genetic
variability than the early population geneticists had assumed in their
models, and that genetically distinct sub-populations were important.
Dobzhansky argued that natural selection worked to maintain genetic
diversity as well as driving change. He was influenced by his exposure
in the 1920s to the work of
Sergei Chetverikov,
who had looked at the role of recessive genes in maintaining a
reservoir of genetic variability in a population before his work was
shut down by the rise of
Lysenkoism in the
Soviet Union.
By 1937, Dobzhansky was able to argue that mutations were the main
source of evolutionary changes and variability, along with chromosome
rearrangements, effects of genes on their neighbours during development,
and polyploidy. Next, genetic drift (he used the term in 1941),
selection, migration, and geographical isolation could change gene
frequencies. Thirdly, mechanisms like ecological or sexual isolation and
hybrid sterility could fix the results of the earlier processes.
Ford's ecological genetics, 1940
E. B. Ford was an experimental naturalist who wanted to test natural selection in nature, virtually inventing the field of
ecological genetics.
His work on natural selection in wild populations of butterflies and
moths was the first to show that predictions made by R. A. Fisher were
correct. In 1940, he was the first to describe and define
genetic polymorphism, and to predict that
human blood group polymorphisms might be maintained in the population by providing some protection against disease. His 1949 book
Mendelism and Evolution helped to persuade Dobzhansky to change the emphasis in the third edition of his famous textbook
Genetics and the Origin of Species from drift to selection.
Schmalhausen's stabilizing selection, 1941
Ivan Schmalhausen developed the theory of
stabilizing selection,
the idea that selection can preserve a trait at some value, publishing a
paper in Russian titled "Stabilizing selection and its place among
factors of evolution" in 1941 and a monograph
Factors of Evolution: The Theory of Stabilizing Selection
in 1945. He developed it from J. M. Baldwin's 1902 concept that changes
induced by the environment will ultimately be replaced by hereditary
changes (including the
Baldwin effect
on behavior), following that theory's implications to their Darwinian
conclusion, and bringing him into conflict with Lysenkoism. Schmalhausen
observed that stabilizing selection would remove most variations from
the norm, most mutations being harmful. Dobzhansky called the work "an important missing link in the modern view of evolution".
Huxley's popularising synthesis, 1942
In 1942,
Julian Huxley's serious but popularizing
Evolution: The Modern Synthesis
introduced a name for the synthesis and intentionally set out to
promote a "synthetic point of view" on the evolutionary process. He
imagined a wide synthesis of many sciences: genetics, developmental
physiology, ecology, systematics, palaeontology, cytology, and
mathematical analysis of biology, and assumed that evolution would
proceed differently in different groups of organisms according to how
their genetic material was organized and their strategies for
reproduction, leading to progressive but varying evolutionary trends. His vision was of an "evolutionary humanism",
with a system of ethics and a meaningful place for "Man" in the world
grounded in a unified theory of evolution which would demonstrate
progress leading to man at its summit. Natural selection was in his view
a "fact of nature capable of verification by observation and
experiment", while the "period of synthesis" of the 1920s and 1930s had
formed a "more unified science", rivaling physics and enabling the "rebirth of Darwinism".
However, the book was not the research text that it appeared to be. In the view of the philosopher of science
Michael Ruse, and in Huxley's own opinion, Huxley was "a generalist, a synthesizer of ideas, rather than a specialist".
Ruse observes that Huxley wrote as if he were adding empirical evidence
to the mathematical framework established by Fisher and the population
geneticists, but that this was not so. Huxley avoided mathematics, for
instance not even mentioning
Fisher's fundamental theorem of natural selection.
Instead, Huxley used a mass of examples to demonstrate that natural
selection is powerful, and that it works on Mendelian genes. The book
was successful in its goal of persuading readers of the reality of
evolution, effectively illustrating topics such as
island biogeography,
speciation, and competition. Huxley further showed that the appearance of long-term
orthogenetic trends – predictable directions for evolution – in the fossil record were readily explained as
allometric growth
(since parts are interconnected). All the same, Huxley did not reject
orthogenesis out of hand, but maintained a belief in progress all his
life, with
Homo sapiens as the end point, and he had since 1912 been influenced by the
vitalist philosopher
Henri Bergson, though in public he maintained an atheistic position on evolution.
Huxley's belief in progress within evolution and evolutionary humanism
was shared in various forms by Dobzhansky, Mayr, Simpson and Stebbins,
all of them writing about "the future of Mankind". Both Huxley and
Dobzhansky admired the palaeontologist priest
Pierre Teilhard de Chardin, Huxley writing the introduction to Teilhard's 1955 book on orthogenesis,
The Phenomenon of Man. This vision required evolution to be seen as the central and guiding principle of biology.
Mayr's allopatric speciation, 1942
Ernst Mayr's key contribution to the synthesis was
Systematics and the Origin of Species, published in 1942.
It asserted the importance of and set out to explain population
variation in evolutionary processes including speciation. He analyzed in
particular the effects of
polytypic species, geographic variation, and isolation by geographic and other means. Mayr emphasized the importance of
allopatric speciation, where geographically isolated sub-populations diverge so far that
reproductive isolation occurs. He was skeptical of the reality of
sympatric speciation
believing that geographical isolation was a prerequisite for building
up intrinsic (reproductive) isolating mechanisms. Mayr also introduced
the
biological species concept
that defined a species as a group of interbreeding or potentially
interbreeding populations that were reproductively isolated from all
other populations. Before he left
Germany for the United States in 1930, Mayr had been influenced by the work of the German biologist
Bernhard Rensch,
who in the 1920s had analyzed the geographic distribution of polytypic
species, paying particular attention to how variations between
populations correlated with factors such as differences in climate.
Simpson's palaeontology, 1944
George Gaylord Simpson was responsible for showing that the modern synthesis was compatible with palaeontology in his 1944 book
Tempo and Mode in Evolution.
Simpson's work was crucial because so many palaeontologists had
disagreed, in some cases vigorously, with the idea that natural
selection was the main mechanism of evolution. It showed that the trends
of linear progression (in for example the
evolution of the horse) that earlier palaeontologists had used as support for
neo-Lamarckism and orthogenesis did not hold up under careful examination. Instead the
fossil record was consistent with the irregular, branching, and non-directional pattern predicted by the modern synthesis.
The Society for the Study of Evolution, 1946
During the
war,
Mayr edited a series of bulletins of the Committee on Common Problems
of Genetics, Paleontology, and Systematics, formed in 1943, reporting on
discussions of a "synthetic attack" on the interdisciplinary problems
of evolution. In 1946, the committee became the Society for the Study of
Evolution, with Mayr, Dobzhansky and Sewall Wright the first of the
signatories. Mayr became the editor of its journal,
Evolution.
From Mayr and Dobzhansky's point of view, suggests the historian of
science Betty Smocovitis, Darwinism was reborn, evolutionary biology was
legitimized, and genetics and evolution were synthesised into a newly
unified science. Everything fitted in to the new framework, except
"heretics" like
Richard Goldschmidt who annoyed Mayr and Dobzhansky by insisting on the possibility of
speciation by macromutation, creating "hopeful monsters". The result was "bitter controversy".
Stebbins's botany, 1950
Definitions by the founders
The
modern synthesis was defined differently by its various founders, with
differing numbers of basic postulates, as shown in the table.
After the synthesis
After the synthesis, evolutionary biology continued to develop with major contributions from workers including W. D. Hamilton, George C. Williams, E. O. Wilson, Edward B. Lewis and others.
Hamilton's inclusive fitness, 1964
In 1964,
W. D. Hamilton published two papers on "The Genetical Evolution of Social Behaviour". These defined
inclusive fitness
as the number of offspring equivalents an individual rears, rescues or
otherwise supports through its behavior. This was contrasted with
personal reproductive fitness, the number of offspring that the
individual directly begets. Hamilton, and others such as
John Maynard Smith,
argued that a gene's success consisted in maximizing the number of
copies of itself, either by begetting them or by indirectly encouraging
begetting by related individuals who shared the gene, the theory of
kin selection.
Williams's gene-centred evolution, 1966
Wilson's sociobiology, 1975
In 1975,
E. O. Wilson published his controversial book
Sociobiology: The New Synthesis, the subtitle alluding to the modern synthesis
as he attempted to bring the study of animal society into the
evolutionary fold. This appeared radically new, although Wilson was
following Darwin, Fisher, Dawkins and others. Critics such as
Gerhard Lenski
noted that he was following Huxley, Simpson and Dobzhansky's approach,
which Lenski considered needlessly reductive as far as human society was
concerned. By 2000, the proposed discipline of
sociobiology had morphed into the relatively well-accepted discipline of
evolutionary psychology.
Lewis's homeotic genes, 1978
In 1977,
recombinant DNA technology enabled biologists to start to explore the genetic control of development. The growth of
evolutionary developmental biology from 1978, when
Edward B. Lewis discovered
homeotic genes, showed that many so-called
toolkit genes
act to regulate development, influencing the expression of other genes.
It also revealed that some of the regulatory genes are extremely
ancient, so that animals as different as insects and mammals share
control mechanisms; for example, the
Pax6 gene is involved in forming the
eyes of mice and of fruit flies. Such
deep homology provided strong evidence for evolution and indicated the paths that evolution had taken.
Later syntheses
In 1982, a historical note on a series of evolutionary biology books
could state without qualification that evolution is the central
organizing principle of biology. Smocovitis commented on this that "What
the architects of the synthesis had worked to construct had by 1982
become a matter of fact", adding in a footnote that "the centrality of
evolution had thus been rendered
tacit knowledge, part of the
received wisdom of the profession".
By the late 20th century, however, the modern synthesis was
showing its age, and fresh syntheses to remedy its defects and fill in
its gaps were proposed from different directions. These have included
such diverse fields as the
study of society, developmental biology, epigenetics,
molecular biology,
microbiology,
genomics,
symbiogenesis, and
horizontal gene transfer. The physiologist
Denis Noble
argues that these additions render neo-Darwinism in the sense of the
early 20th century's modern synthesis "at the least, incomplete as a
theory of evolution", and one that has been falsified by later biological research.
Michael Rose and Todd Oakley note that evolutionary biology, formerly divided and "
Balkanized",
has been brought together by genomics. It has in their view discarded
at least five common assumptions from the modern synthesis, namely that
the genome is always a well-organized set of genes; that each gene has a
single function; that species are well adapted biochemically to their
ecological niches; that species are the durable units of evolution, and
all levels from organism to organ, cell and molecule within the species
are characteristic of it; and that the design of every organism and
cell is efficient. They argue that the "new biology" integrates
genomics,
bioinformatics, and evolutionary genetics into a general-purpose toolkit for a "Postmodern Synthesis".
Pigliucci's extended evolutionary synthesis, 2007
Koonin's 'post-modern' evolutionary synthesis, 2009
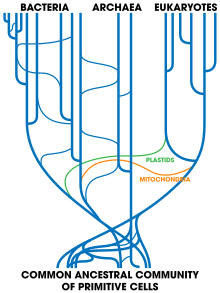
In 2009, Darwin's 200th anniversary, the
Origin of Species' 150th, and the 200th of Lamarck's "early evolutionary synthesis",
Philosophie Zoologique, the evolutionary biologist
Eugene Koonin stated that while "the edifice of the [early 20th century] Modern Synthesis has crumbled, apparently, beyond repair",
a new 21st century synthesis could be glimpsed. Three interlocking
revolutions had, he argued, taken place in evolutionary biology:
molecular, microbiological, and genomic. The
molecular revolution
included the neutral theory, that most mutations are neutral and that
purifying selection happens more often than the positive form, and that
all current life evolved from
a single common ancestor. In microbiology, the synthesis has expanded to cover the
prokaryotes, using
ribosomal RNA to form a
tree of life. Finally,
genomics
brought together the molecular and microbiological syntheses, noting
that a molecular view shows that the tree of life is problematic. In
particular,
horizontal gene transfer between
bacteria means that prokaryotes freely share genes, challenging Mayr's foundational definition of
species. Further, horizontal gene transfer, gene duplication, and "momentous events" like
endosymbiosis
enable evolution to proceed in sudden jumps, ending the old
gradualist-saltationist debate by showing that on this point Darwin's
gradualism was wrong. The idea of
progress in biology, too, is seen to be wrong, along with the modern synthesis belief in
pan-adaptationism, that everything is optimally adapted: genomes plainly are not. Many of these points had already been made by other researchers such as Ulrich Kutschera and
Karl J. Niklas.
Towards a replacement synthesis
Inputs
to the modern synthesis, with other topics (inverted colours) such as
developmental biology that were not joined with evolutionary biology
until the turn of the 21st century
Biologists, alongside scholars of the history and philosophy of
biology, have continued to debate the need for, and possible nature of, a
replacement synthesis. For example, in 2017 Philippe Huneman and Denis
M. Walsh stated in their book
Challenging the Modern Synthesis
that numerous theorists had pointed out that the disciplines of
embryological developmental theory, morphology, and ecology had been
omitted. They noted that all such arguments amounted to a continuing
desire to replace the modern synthesis with one that united "all
biological fields of research related to evolution, adaptation, and
diversity in a single theoretical frame."
They observed further that there are two groups of challenges to the
way the modern synthesis viewed inheritance. The first is that other
modes such as
epigenetic inheritance,
phenotypic plasticity, the
Baldwin effect, and the
maternal effect
allow new characteristics to arise and be passed on, and for the genes
to catch up with the new adaptations later. The second is that all such
mechanisms are part, not of an inheritance system, but a
developmental system:
the fundamental unit is not a discrete selfishly competing gene, but a
collaborating system that works at all levels from genes and cells to
organisms and cultures to guide evolution.
Historiography
Looking back at the conflicting accounts of the modern synthesis, the historian Betty Smocovitis notes in her 1996 book Unifying Biology: The Evolutionary Synthesis and Evolutionary Biology
that both historians and philosophers of biology have attempted to
grasp its scientific meaning, but have found it "a moving target"; the only thing they agreed on was that it was a historical event.
In her words "by the late 1980s the notoriety of the evolutionary
synthesis was recognized . . . So notorious did 'the synthesis' become,
that few serious historically minded analysts would touch the subject,
let alone know where to begin to sort through the interpretive mess left
behind by the numerous critics and commentators".


















