| Cerebral cortex | |
|---|---|

Tissue
slice from the brain of an adult macaque monkey (Macaca mulatta). The
cerebral cortex is the outer layer depicted in dark violet. Source:
BrainMaps.org
| |

Golgi-stained neurons in the cortex
| |
| Details | |
| Part of | Cerebrum |
| Identifiers | |
| Latin | Cortex cerebri |
| MeSH | D002540 |
| NeuroNames | 39 |
| NeuroLex ID | birnlex_1494 |
| TA | A14.1.09.003 A14.1.09.301 |
| FMA | 61830 |
The cerebral cortex is the largest region of the cerebrum in the mammalian brain and plays a key role in memory, attention, perception, cognition, awareness, thought, language, and consciousness. The cerebral cortex is the most anterior (rostral) brain region and consists of an outer zone of neural tissue called gray matter, which contains neuronal cell bodies. It is also divided into left and right cerebral hemispheres by the longitudinal fissure, but the two hemispheres are joined at the midline by the corpus callosum.
At the cellular and circuit level, the cerebral cortex is characterized by two primary organizational features:
- across its surface it is divided into functional areas that serve various sensory, motor, and cognitive functions, and
- it is subdivided into several layers that organize the input and output connectivity of resident neurons.
In large mammals, the cerebral cortex is usually folded, providing a greater surface area in the confined volume of the cranium. Increased surface area is thought to be important because it allows for the addition and evolution of a greater diversity of functional modules, or areas. A fold or ridge in the cortex is termed a gyrus (plural gyri) and a groove is termed a sulcus (plural sulci). These surface convolutions appear during fetal development and continue to mature after birth through the process of gyrification. In the human brain the majority of the cerebral cortex is not visible from the outside, but buried in the sulci.
The cerebral cortex contains a large number of neuronal and glial cell bodies, as well as their intricate dendritic formations and axonal projections, which connect at synapses to form basic functional circuits. The cerebral cortex is entirely made of gray matter, contrasting with the underlying white matter, which consists mainly of axons traveling to and from the cortex, their myelinated sheaths, and the cell bodies of oligodendrocytes.
Structure
Layers

Cerebral
cortex. (Poirier.) To the left, the groups of cells; to the right, the
systems of fibers. Quite to the left of the figure a sensory nerve fiber
is shown. Cell body layers are labeled on the left, and fiber layers
are labeled on the right.

Three drawings of cortical lamination by Santiago Ramon y Cajal, each showing a vertical cross-section, with the surface of the cortex at the top. Left: Nissl-stained visual cortex of a human adult. Middle: Nissl-stained motor cortex of a human adult. Right: Golgi-stained cortex of a 1 1⁄2 month old infant. The Nissl stain shows the cell bodies of neurons; the Golgi stain shows the dendrites and axons of a random subset of neurons.

Micrograph showing the visual cortex (predominantly pink). Subcortical white matter (predominantly blue) is seen at the bottom of the image. HE-LFB stain.
Staining cross-sections of the cortex to reveal the position of neuronal cell bodies and the intracortical axon tracts allowed neuroanatomists in the early 20th century to produce a detailed description of the laminar structure of the cortex in different species. After the work of Korbinian Brodmann (1909) the neurons of the cerebral cortex are grouped into six main layers, from outside (pial surface) to inside (white matter):
- Layer I, the molecular layer, contains few scattered neurons and consists mainly of extensions of apical dendritic tufts of pyramidal neurons and horizontally oriented axons, as well as glial cells. During development Cajal-Retzius and subpial granular layer cells are present in this layer. Also, some spiny stellate cells can be found here. Inputs to the apical tufts are thought to be crucial for the feedback interactions in the cerebral cortex involved in associative learning and attention. While it was once thought that the input to layer I came from the cortex itself, it is now realized that layer I across the cerebral cortex mantle receives substantial input from matrix or M-type thalamus cells (in contrast to core or C-type that go to layer IV).
- Layer II, the external granular layer, contains small pyramidal neurons and numerous stellate neurons.
- Layer III, the external pyramidal layer, contains predominantly small and medium-size pyramidal neurons, as well as non-pyramidal neurons with vertically oriented intracortical axons; layers I through III are the main target of interhemispheric corticocortical afferents, and layer III is the principal source of corticocortical efferents.
- Layer IV, the internal granular layer, contains different types of stellate and pyramidal neurons, and is the main target of thalamocortical afferents from thalamus type C neurons as well as intra-hemispheric corticocortical afferents. The layers above layer IV are also referred to as supragranular layers (layers I-III), whereas the layers below are referred to as infragranular layers (layers V and VI).
- Layer V, the internal pyramidal layer, contains large pyramidal neurons which give rise to axons leaving the cortex and running down to subcortical structures (such as the basal ganglia). In the primary motor cortex of the frontal lobe, layer V contains Betz cells, whose axons travel through the internal capsule, the brain stem and the spinal cord forming the corticospinal tract, which is the main pathway for voluntary motor control.
- Layer VI, the polymorphic or multiform layer, contains few large pyramidal neurons and many small spindle-like pyramidal and multiform neurons; layer VI sends efferent fibers to the thalamus, establishing a very precise reciprocal interconnection between the cortex and the thalamus. That is, layer VI neurons from one cortical column connect with thalamus neurons that provide input to the same cortical column. These connections are both excitatory and inhibitory. Neurons send excitatory fibers to neurons in the thalamus and also send collaterals to the thalamic reticular nucleus that inhibit these same thalamus neurons or ones adjacent to them. One theory is that because the inhibitory output is reduced by cholinergic input to the cerebral cortex, this provides the brainstem with adjustable "gain control for the relay of lemniscal inputs".
Cortical areas that lack a layer IV are called agranular. Cortical areas that have only a rudimentary layer IV are called dysgranular. Information processing within each layer is determined by different temporal dynamics with that in the layers II/III having a slow 2 Hz oscillation while that in layer V having a fast 10–15 Hz one.
Areas
Based on the differences in lamination the cerebral cortex can be classified into two parts, the large area of neocortex and the much smaller area of allocortex:- The neocortex (also known as the isocortex or neopallium) is the part of the mature cerebral cortex with six distinct layers. Examples of neocortical areas include the granular primary motor cortex, also known as Brodmann area 4, and the striate primary visual cortex, or Brodmann area 17. The neocortex has two types of cortices, the true isocortex and the proisocortex. The proisocortex contains Brodmann areas 24, 25, and 32
- The allocortex is the part of the cerebral cortex with less than six layers and has three regions, the paleocortex with three cortical laminae and the archicortex which has four or five, and a transitional area adjacent to the allocortex, the periallocortex. Examples of allocortex are the olfactory cortex and the hippocampus.

 Lateral surface of the cerebral cortex
Lateral surface of the cerebral cortex Medial surface of the cerebral cortex
Medial surface of the cerebral cortex
Brodmann areas
Different parts of the cerebral cortex are involved in different cognitive and behavioral functions. The differences show up in a number of ways: the effects of localized brain damage, regional activity patterns exposed when the brain is examined using functional imaging techniques, connectivity with subcortical areas, and regional differences in the cellular architecture of the cortex. Neuroscientists describe most of the cortex—the part they call the neocortex—as having six layers, but not all layers are apparent in all areas, and even when a layer is present, its thickness and cellular organization may vary. Scientists have constructed maps of cortical areas on the basis of variations in the appearance of the layers as seen with a microscope. One of the most widely used schemes came from Korbinian Brodmann, who split the cortex into 51 different areas and assigned each a number (many of these Brodmann areas have since been subdivided). For example, Brodmann area 1 is the primary somatosensory cortex, Brodmann area 17 is the primary visual cortex, and Brodmann area 25 is the anterior cingulate cortex.Thickness
For mammals, species with larger brains (in absolute terms, not just in relation to body size) tend to have thicker cortices. The range, however, is not very great; only a factor of 7 differentiates between the thickest and thinnest cortices. The smallest mammals, such as shrews, have a neocortical thickness of about 0.5 mm; the ones with the largest brains, such as humans and fin whales, have thicknesses of 2.3–2.8 mm. There is an approximately logarithmic relationship between brain weight and cortical thickness.
Cortical blood supply
Magnetic resonance imaging of the brain (MRI) makes it possible to get a measure for the thickness of the human cerebral cortex and relate it to other measures. The thickness of different cortical areas varies but in general, sensory cortex is thinner than motor cortex. One study has found some positive association between the cortical thickness and intelligence. Another study has found that the somatosensory cortex is thicker in migraine sufferers, though it is not known if this is the result of migraine attacks or the cause of them. A later study using a larger patient population reports no change in the cortical thickness in migraine sufferers. A genetic disorder of the cerebral cortex, whereby decreased folding in certain areas results in a microgyrus, where there are four layers instead of six, is in some instances seen to be related to dyslexia.
Folds
The cerebral cortex is folded in a way that allows a large surface area to fit within the confines of the skull. When unfolded, each cerebral hemisphere cortex has a total surface area of about 1.3 square feet (0.12 m2). The folding is inward away from the surface of the brain, and is also present on the medial surface of the brain within the longitudinal fissure. The folding creates a looping, meandering, 'worm-pile' appearance of the surface of the brain. The peaks of folds are called gyri (singular gyrus), and the valleys are called sulci (singular sulcus).Development

Human cortical development between 26 and 39 week gestational age
The ontogenic development of the cerebral cortex is a complex and finely tuned process influenced by the interplay between genes and environment. The cerebral cortex develops from the most anterior part of the neural plate, a specialized part of the embryonic ectoderm. The neural plate folds and closes to form the neural tube. From the cavity inside the neural tube develops the ventricular system, and, from the epithelial cells of its walls, the neurons and glia of the nervous system. The most anterior (front, or cranial) part of the neural plate, the prosencephalon, which is evident before neurulation begins, gives rise to the cerebral hemispheres and its later cortex.
Cortical neurons are generated within the ventricular zone, next to the ventricles. At first, this zone contains progenitor cells, which divide to produce glial cells and neurons. The glial fibers produced in the first divisions of the progenitor cells are radially oriented, spanning the thickness of the cortex from the ventricular zone to the outer, pial surface, and provide scaffolding for the migration of neurons outwards from the ventricular zone. The first divisions of the progenitor cells are symmetric, which duplicates the total number of progenitor cells at each mitotic cycle. Then, some progenitor cells begin to divide asymmetrically, producing one postmitotic cell that migrates along the radial glial fibers, leaving the ventricular zone, and one progenitor cell, which continues to divide until the end of development, when it differentiates into a glial cell or an ependymal cell. As the G1 phase of mitosis is elongated, in what is seen as selective cell-cycle lengthening, the newly-born neurons migrate to more superficial layers of the cortex. The migrating daughter cells become the pyramidal cells of the cerebral cortex. The development process is time ordered and regulated by hundreds of genes and epigenetic regulatory mechanisms.
The layered structure of the mature cerebral cortex is formed during development. The first pyramidal neurons generated migrate out of the ventricular zone and subventricular zone, together with reelin producing Cajal–Retzius neurons, from the preplate. Next, a cohort of neurons migrating into the middle of the preplate divides this transient layer into the superficial marginal zone, which will become layer one of the mature neocortex, and the subplate, forming a middle layer called the cortical plate. These cells will form the deep layers of the mature cortex, layers five and six. Later born neurons migrate radially into the cortical plate past the deep layer neurons, and become the upper layers (two to four). Thus, the layers of the cortex are created in an inside-out order. The only exception to this inside-out sequence of neurogenesis occurs in the layer I of primates, in which, in contrast to rodents, neurogenesis continues throughout the entire period of corticogenesis.

Depicted
in blue, Emx2 is highly expressed at the caudomedial pole and
dissipates outward. Pax6 expression is represented in purple and is
highly expressed at the rostral lateral pole. (Adapted from Sanes, D.,
Reh, T., & Harris, W. (2012). Development of the Nervous System (3rd ed.). Burlington: Elsevier Science)
The map of functional cortical areas, which include primary motor and visual cortex, originates from a 'protomap', which is regulated by molecular signals such as fibroblast growth factor FGF8 early in embryonic development. These signals regulate the size, shape, and position of cortical areas on the surface of the cortical primordium, in part by regulating gradients of transcription factor expression, through a process called cortical patterning. Examples of such transcription factors include the genes EMX2 and PAX6. Together, both transcription factors form an opposing gradient of expression. Pax6 is highly expressed at the rostral lateral pole, while Emx2 is highly expressed in the caudomedial pole. The establishment of this gradient is important for proper development. For example, mutations in Pax6 can cause expression levels of Emx2 to expand out of its normal expression domain, which would ultimately lead to an expansion of the areas normally derived from the caudal medial cortex, such as the visual cortex. On the contrary, if mutations in Emx2 occur, it can cause the Pax6-expressing domain to expand and result in the frontal and motor cortical regions enlarging. Therefore, researchers believe that similar gradients and signaling centers next to the cortex could contribute to the regional expression of these transcription factors.
Two very well studied patterning signals for the cortex include FGF and retinoic acid. If FGFs are misexpressed in different areas of the developing cortex, cortical patterning is disrupted. Specifically, when Fgf8 is increased in the anterior pole, Emx2 is downregulated and a caudal shift in the cortical region occurs. This ultimately causes an expansion of the rostral regions. Therefore, Fgf8 and other FGFs play a role in the regulation of expression of Emx2 and Pax6 and represent how the cerebral cortex can become specialized for different functions.
Rapid expansion of the cortical surface area is regulated by the amount of self-renewal of radial glial cells and is partly regulated by FGF and Notch genes. During the period of cortical neurogenesis and layer formation, many higher mammals begin the process of gyrification, which generates the characteristic folds of the cerebral cortex. Gyrification is regulated by a DNA-associated protein Trnp1 and by FGF and SHH signaling.

Neurogenesis is shown in red and lamination is shown in blue. Adapted from (Sur et al. 2001)
The cerebral cortex is composed of a heterogenous population of cells that give rise to different cell types. The majority of these cells are derived from radial glia migration that form the different cell types of the neocortex and it is a period associated with an increase in neurogenesis. Similarly, the process of neurogenesis regulates lamination to form the different layers of the cortex. During this process there is an increase in the restriction of cell fate that begins with earlier progenitors giving rise to any cell type in the cortex and later progenitors giving rise only to neurons of superficial layers. This differential cell fate creates an inside-out topography in the cortex with younger neurons in superficial layers and older neurons in deeper layers. In addition, laminar neurons are stopped in S or G2 phase in order to give a fine distinction between the different cortical layers. Laminar differentiation is not fully complete until after birth since during development laminar neurons are still sensitive to extrinsic signals and environmental cues.
Although the majority of the cells that compose the cortex are derived locally from radial glia there is a subset population of neurons that migrate from other regions. Radial glia give rise to neurons that are pyramidal in shape and use glutamate as a neurotransmitter, however these migrating cells contribute neurons that are stellate-shaped and use GABA as their main neurotransmitter. These GABAergic neurons are generated by progenitor cells in the medial ganglionic eminence (MGE) that migrate tangentially to the cortex via the subventricular zone. This migration of GABAergic neurons is particularly important since GABA receptors are excitatory during development. This excitation is primarily driven by the flux of chloride ions through the GABA receptor, however in adults chloride concentrations shift causing an inward flux of chloride that hyperpolarizes postsynaptic neurons.
Evolution
Of all the different brain regions, the cerebral cortex shows the largest evolutionary variation and has evolved most recently. In contrast to the highly conserved circuitry of the medulla oblongata, for example, which serves critical functions such as regulation of heart and respiration rates, many areas of the cerebral cortex are not strictly necessary for survival. Thus, the evolution of the cerebral cortex has seen the advent and modification of new functional areas—particularly association areas that do not directly receive input from outside the cortex.A key theory of cortical evolution is embodied in the radial unit hypothesis and related protomap hypothesis, first proposed by Rakic. This theory states that new cortical areas are formed by the addition of new radial units, which is accomplished at the stem cell level. The protomap hypothesis states that the cellular and molecular identity and characteristics of neurons in each cortical area are specified by cortical stem cells, known as radial glial cells, in a primordial map. This map is controlled by secreted signaling proteins and downstream transcription factors.
Function
Connections
The cerebral cortex is connected to various subcortical structures such as the thalamus and the basal ganglia, sending information to them along efferent connections and receiving information from them via afferent connections. Most sensory information is routed to the cerebral cortex via the thalamus. Olfactory information, however, passes through the olfactory bulb to the olfactory cortex (piriform cortex). The majority of connections are from one area of the cortex to another, rather than from subcortical areas; Braitenberg and Schüz (1998) claim that in primary sensory areas, at the cortical level where the input fibres terminate, up to 20% of the synapses are supplied by extracortical afferents but that in other areas and other layers the percentage is likely to be much lower.Cortical areas

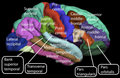
Lateral surface of the human cerebral cortex

Medial surface of the human cerebral cortex
Sensory areas
The sensory areas are the cortical areas that receive and process information from the senses. Parts of the cortex that receive sensory inputs from the thalamus are called primary sensory areas. The senses of vision, audition, and touch are served by the primary visual cortex, primary auditory cortex and primary somatosensory cortex respectively. In general, the two hemispheres receive information from the opposite (contralateral) side of the body. For example, the right primary somatosensory cortex receives information from the left limbs, and the right visual cortex receives information from the left visual field. The organization of sensory maps in the cortex reflects that of the corresponding sensing organ, in what is known as a topographic map. Neighboring points in the primary visual cortex, for example, correspond to neighboring points in the retina. This topographic map is called a retinotopic map. In the same way, there exists a tonotopic map in the primary auditory cortex and a somatotopic map in the primary sensory cortex. This last topographic map of the body onto the posterior central gyrus has been illustrated as a deformed human representation, the somatosensory homunculus, where the size of different body parts reflects the relative density of their innervation. Areas with lots of sensory innervation, such as the fingertips and the lips, require more cortical area to process finer sensation.Motor areas
The motor areas are located in both hemispheres of the cortex. They are shaped like a pair of headphones stretching from ear to ear. The motor areas are very closely related to the control of voluntary movements, especially fine fragmented movements performed by the hand. The right half of the motor area controls the left side of the body, and vice versa.Two areas of the cortex are commonly referred to as motor:
- Primary motor cortex, which executes voluntary movements
- Supplementary motor areas and premotor cortex, which select voluntary movements.
- Posterior parietal cortex, which guides voluntary movements in space
- Dorsolateral prefrontal cortex, which decides which voluntary movements to make according to higher-order instructions, rules, and self-generated thoughts.
Association areas
The association areas are the parts of the cerebral cortex that do not belong to the primary regions. They function to produce a meaningful perceptual experience of the world, enable us to interact effectively, and support abstract thinking and language. The parietal, temporal, and occipital lobes - all located in the posterior part of the cortex - integrate sensory information and information stored in memory. The frontal lobe or prefrontal association complex is involved in planning actions and movement, as well as abstract thought. Globally, the association areas are organized as distributed networks. Each network connects areas distributed across widely spaced regions of the cortex. Distinct networks are positioned adjacent to one another yielding a complex series of interwoven networks. The specific organization of the association networks is debated with evidence for interactions, hierarchical relationships, and competition between networks. In humans, association networks are particularly important to language function. In the past it was theorized that language abilities are localized in the left hemisphere in areas 44/45, the Broca's area, for language expression and area 22, the Wernicke's area, for language reception. However, language is no longer limited to easily identifiable areas. More recent research suggests that the processes of language expression and reception occur in areas other than just those structures around the lateral sulcus, including the frontal lobe, basal ganglia, cerebellum, and pons.Clinical significance

There
is marked cortical atrophy in Alzheimer's disease, associated with loss
of gyri and sulci in the temporal lobe and parietal lobe, and parts of
the frontal cortex and cingulate gyrus.
Neurodegenerative diseases such as Alzheimer's disease and Lafora disease, show as a marker, an atrophy of the grey matter of the cerebral cortex.
There are many neurodevelopmental abnormalities that can lead to a wide variety of behavioral and cognitive deficits. There are several situations in development in which both intrinsic and extrinsic factors can highly influence the course of nervous system formation. One very prominent intrinsic factor (random gene mutation) has given rise to many different classes of neurodevelopmental disorders. For example, Fragile X-Syndrome is a neurodevelopmental disease characterized by poor eye contact with others, an extreme aversion to physical/social contact, and obsessive repetition in behavioral patterns. This is an X-linked chromosomal disorder in which the FMR1 gene is found to have nearly 200 copies, instead of its intended 30. This causes the gene to become heavily methylated, which subsequently turns off expression of FMR1. Efficient functioning of this gene is known to play a role in localized protein synthesis at dendritic spines, which is essential for proper synaptogenesis and learning and memory function.

Maternal alcohol consumption leads to death of neural crest cells which are the building blocks of certain neural tissues.
Another primary example of intrinsic neurodevelopmental deficits is Rett Syndrome, which is an X-linked single gene mutation characterized by a loss of speech and hand coordination, intellectual regression and progressive loss of motor control. This disorder is thought to arise from a mutation in the MeCP2 gene, which encodes for a transcription factor associated with chromatin remodeling. Mutations in this gene have been linked to a decreased expression of the gene that codes for BDNF (brain-derived neurotrophic factor), which is a common gene used in neurodevelopment.
Apart from intrinsic gene mutation, there are many extrinsic (environmental) factors that can greatly influence the neurodevelopmental process. One of the most commonly studied factors is the effect of maternal alcohol consumption during gestation. Infants born with defects from this factor are said to have Fetal Alcohol Spectrum Disorder (FASD). FASD is used to describe a collection of developmental disorders associated with the consumption of alcohol whilst pregnant. The spectrum includes individuals with both extreme and mild FASD which present with a variety of neurocognitive and neurobehavioral deficits. This includes deficits is sustained and focused attention, learning and memory problems, verbal processing problems, receptive language difficulties, hyperactivity, as well as a range of psychiatric impairments. Apoptotic cell death caused by an excess of glutamate activity and GABA withdrawal is thought to play a role in the loss of neuronal fibers critical for normal brain development. In addition to neuronal death, there is evidence suggesting that an improper formation of interneuronal connections and cell adhesion molecule malformation are also associated with FASD. Improper neuronal migration and synaptogenesis have been shown to occur is several cases of FASD, which could explain the extreme deficits in learning and memory.

Individuals
with Fetal Alcohol Spectrum Disorder (FASD) exhibit a range of
cognitive and behavioral deficits that relate heavily to neural
pathologies associated with maternal alcohol consumption.
Other animals
The cerebral cortex is derived from the pallium, a layered structure found in the forebrain of all vertebrates. The basic form of the pallium is a cylindrical layer enclosing fluid-filled ventricles. Around the circumference of the cylinder are four zones, the dorsal pallium, medial pallium, ventral pallium, and lateral pallium, which are thought respectively to give rise to the neocortex, hippocampus, amygdala, and olfactory cortex.Until recently no counterpart to the cerebral cortex had been recognized in invertebrates. However, a study published in the journal Cell in 2010, based on gene expression profiles, reported strong affinities between the cerebral cortex and the mushroom bodies of the ragworm Platynereis dumerilii. Mushroom bodies are structures in the brains of many types of worms and arthropods that are known to play important roles in learning and memory; the genetic evidence indicates a common evolutionary origin, and therefore indicates that the origins of the earliest precursors of the cerebral cortex date back to the early Precambrian era.
Additional images
 Motor and sensory regions of the cerebral cortex
Motor and sensory regions of the cerebral cortex Motor and sensory regions of the cerebral cortex
Motor and sensory regions of the cerebral cortex Cortical areas
Cortical areas





